Слухова зона кори головного мозку розташована в

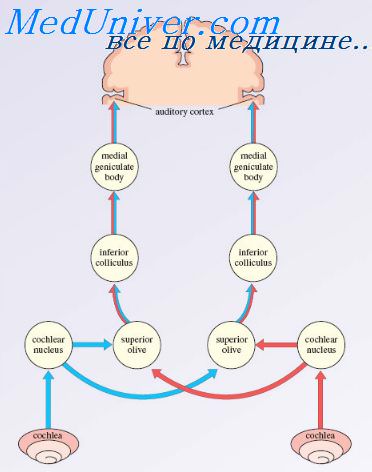
Слуховая кора. Слуховая функция коры головного мозгаНа рисунке показана проекционная слуховая зона мозговой коры, которая лежит главным образом в надвисочной плоскости верхней височной доли, но также распространяется на латеральную сторону височной доли, на большую часть островковой коры и даже на латеральную часть теменной покрышки. На рисунке показаны две отдельные части слуховой коры: первичная слуховая кора и ассоциативная слуховая кора (называемая также вторичной слуховой корой). Первичная слуховая кора непосредственно возбуждается проекциями от медиального коленчатого тела, тогда как ассоциативная слуховая кора возбуждается вторично импульсами из первичной слуховой коры, а также проекциями из таламических ассоциативных зон, прилежащих к медиальному коленчатому телу. Восприятие частоты звука в первичной слуховой коре. В первичной слуховой коре и ассоциативной слуховой коре обнаружены, по крайней мере, шесть тонотопических карт. В каждой из этих карт высокочастотные звуки возбуждают нейроны у одного конца карты, а низкочастотные звуки — у противоположного конца. В основном, низкочастотные звуки локализуются спереди, а высокочастотные — сзади.
Это справедливо не для всех карт. Возникает вопрос: почему слуховая кора имеет так много разных карт? Ответ, вероятно, в том, что каждая из отдельных областей анализирует одно из специфических свойств звука. Например, одна из больших карт в первичной слуховой коре, вероятно, различает сами звуковые частоты и дает человеку физическое ощущение высоты звуков. Другая карта, вероятно, используется для определения направления, откуда исходит звук. Другие области слуховой коры выделяют особые качества, например внезапное начало звуков, или, возможно, особые модуляции, например выделение звуков определенной частоты из шума. Диапазон частот, на которые реагирует каждый нейрон слуховой коры, гораздо уже, чем в улитке и релейных ядрах мозгового ствола. Вновь обратившись к рисунку, можно видеть, что базальная мембрана улитки стимулируется звуками всех частот, и такое же широкое звуковое представительство обнаруживается в улитковых ядрах. Однако когда возбуждение достигает мозговой коры, большинство звукочувствительных нейронов реагируют лишь на узкий, а не на широкий диапазон частот. Следовательно, где-то по пути механизмы анализа «обостряют» реакцию на частоту. Полагают, что причиной этого обостряющего эффекта является главным образом феномен латерального торможения. Это значит, что стимуляция улитки одной частотой тормозит звуковые частоты с обеих сторон этой первичной частоты; причиной являются коллатеральные волокна, ответвляющиеся от первичного сигнального пути и оказывающие тормозное влияние на прилежащие пути. Важность такого эффекта показана также для усиления особенностей соматосенсорных, зрительных и других типов ощущений. Многие нейроны слуховой коры, особенно в ассоциативной слуховой коре, не просто реагируют на специфические звуковые частоты в ухе. Полагают, что эти нейроны «связывают» разные звуковые частоты друг с другом или звуковую информацию с информацией от других сенсорных областей коры. Действительно, теменная часть ассоциативной слуховой коры частично перекрывает соматосенсорную область II, что, вероятно, обеспечивает возможность ассоциации слуховой информации с соматосенсорной. – Также рекомендуем “Различение звуковых образов. Определение направления звука” Оглавление темы “Физиология слуховой системы”: |
Источник
Оглавление темы “Физиология проводящего пути слуха.”:
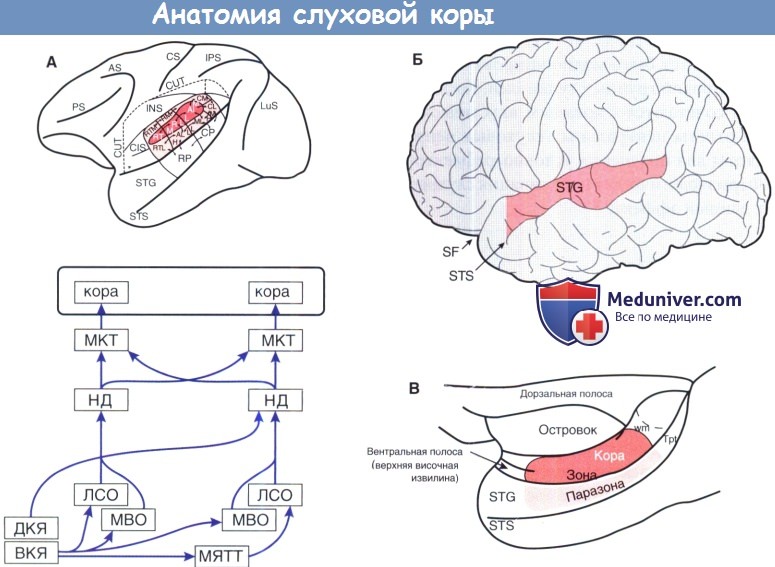
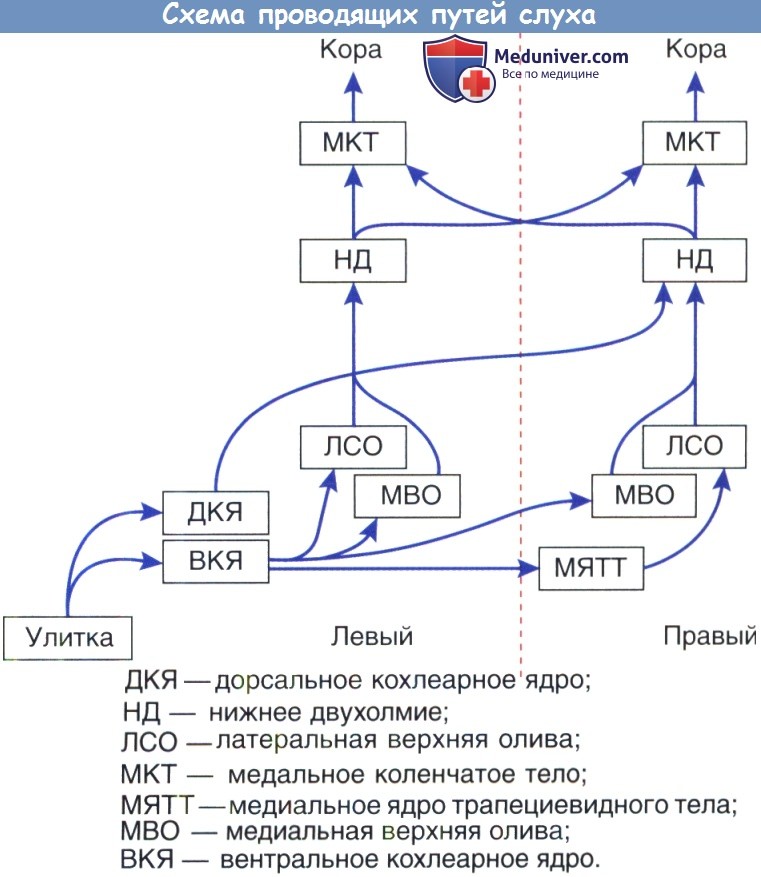
Анатомия слуховой корыСлуховая кора получает все входящие звуковые представления от таламуса. В то время как отдельные подкорковые структуры отвечают за извлечение специфических особенностей из акустических стимулов, слуховая кора должна рекомбинировать эти параллельные потоки информации в интегрированные звуковые восприятия. Эти разнообразные восприятия включают в себя распознавание источников звука, локализацию источника звука, распознавание голосов, интерпретации звуков в биологическом контексте, слуховую память и достигаемую тренировками пластичность. Большая часть наших знаний, относящихся к структуре и функции слуховой коры, была получена в исследованиях на различных видах животных, при этом некоторые из самых интригующих находок получены в работах с приматами. Учитывая ограниченность использования экспериментальных животных в качестве системной модели для изучения высших когнитивных процессов, наши знания о функциях человека значительно продвинулись благодаря косвенным измерениям нервной деятельности, которые могут быть применены к людям, в частности, функциональной нейровизуализации. Технические трудности изучения слуховой функции человека в естественных условиях осложняются выраженной разнохарактерностью слуховой коры с ее многочисленными регионами и полями, каждое из которых обладает своими собственными отличительными анатомическими и физиологическими свойствами. Было использовано много различных анатомических подходов для определения подразделов слуховой коры, в том числе цитоархитектонические и хемоархитектонические методы, в дополнение к изучению нервных связей, участвующих в таламокортикальных, кортико-кортикальных и кортико-фугальных связях. Также были использованы физиологические исследования, основанные на свойствах рецепторного поля, определения НЧ (или частотной характеристики), спектральной полосы целевых нейронов с учетом качества временного ответа. В результате этих исследований четко установлено, что первичное слуховое поле, часто упоминающееся как А1, существует почти во всех видах, включая человека. За А1, однако, было трудно установить единые анатомо-функциональные критерии, которые действительны для разных видов или даже разных индивидуумов одного и того же вида. Анатомия слуховой коры. А1 составляет первую стадию корковой обработки звука. Большая часть того, что нам известно об анатомии А1, основано на исследовании приматов, особенно обезьян, и поэтому, на данный момент, некоторые невыясненные детали у людей могут отличаться. На гистологическом уровне у А1 есть несколько характерных особенностей кониокортекса, включая плотно упакованные маленькие клетки в четвертом слое, обильно миелинизированные волокна и наличие цитохромоксидазы. Как указано ранее, А1 получает большую часть входящих звуков непосредственно от вентральной части таламуса. Функционально, нейроны А1 имеют тенденцию отвечать на чистые тоны хорошо с хорошо настроенными диапазонами НЧ и тонотопическими проведением изочастотного диапазона. Hackett и Kaas используют термин ядро для описания первичных или первично-подобных регионов, которые включают в себя три области (А1, ростральные или Р, и ростротемпоральные или РТ). Эти области представляются активируемыми параллельно, и каждая из этих основных зон демонстрирует отличительные нейронные соединения с другими участками коры. Эти три области организованы каудо-рострально по плоскости боковой щели и имеют тонотопическое расположение. В организме человека большая часть извилины Гешля (поперечная височная извилина) составляет слуховое ядро. Цитоархитектоническим отличием области зоны слуховой коры является то, что она окружает централизованное ядро. Эта область получает несколько входов от вентрального таламического ядра (ВТЯ), его основные таламические входы происходят в дорсальном (ДТЯ), и медиальном ядрах (МТЯ). Нейроны в области зоны получают почти все свои слуховые входы из основных областей и не показывают предварительную активность. Третья область паразоны, точно вентрально к зоне, получает плотные связи от зоны, но почти ни одной от ядра. Паразона не получает таламические входы от дорсального таламического ядра (ДТЯ) и медиального таламического ядра (МТЯ), и корковые входы неслуховых областей прилежат к верхней височной борозде. Эта схема может играть важную роль в полисенсорной обработке (аудиовизуальные взаимодействия). Дополнительные префронтальные связи играют определенную роль в слуховой памяти и зрительно-пространственной ориентации.
Из областей паразоны и зоны звуковые сигналы поступают на четвертый уровень невральной обработки в пределах височных, париетальных, и лобных долей. Смысл этих отношений заключается в том, что многие участки мозга, даже те, что строго не считаются центрами обработки звуков, получают звуковые импульсы и имеют решающее значение для их правильной обработки. Полисенсорные взаимодействия здесь имеют решающее значение, и необходимы дальнейшие исследования для выяснения точной причины нервных взаимодействий между слуховыми и зрительными участками коры. Кроме того, роль гомологичных областей мозга, связанных с мозолистым телом, очень плохо изучена. Несмотря на это, передача звуковой информации происходит именно восходящим образом, что в конечном итоге позволяет людям осознанное восприятие звукового сигнала. Хотя описания слухового ядра, областей зоны и паразоны у обезьян достаточно понятны, наше понимание человеческой слуховой системы остается относительно неполным, принимая во внимание, что большая часть современных знаний получена из недавних исследований с функциональной визуализацией. В частности, это ограниченное понимание может вызывать сложности при интерпретации результатов исследований поражения слуха у людей. Четко установлено, что слуховая кора у людей концентрируется вдоль верхней височной извилины. Верхняя височная плоскость, включающая плоскость полюса височной извилины, извилину Гешля, височную поверхность (спереди назад) — по-видимому, и есть местоположение слуховой коры человека. Эта область имеет особенно высокую вариабельность от человека к человеку, и разделение коры на поля Бродмана может быть затруднено. В 41-м поле по Бродману, что соответствует передней поперечной височной области (где находится зона А1), по-видимому, присутствует вертикальная столбчатая организация слуховых нейронов (именуемые Tel. 1, Tel.О. и Те1.2 медиальнее или кзади к боковым). Как уточняется в первой половине этой статьи, значительный объем обработки слуховой информации происходит в стволе мозга. Таким образом, слуховая кора получает «предварительные» сигналы, которые затем обрабатываются дальше в ядре, регионах зоны и паразоны. А1, по-видимому, участвует в идентификации звуковых объектов, а также интеграции спектральных и временных свойств звуков. В целом наше понимание не-А1 областей является более скудным по сравнению с А1, в частности, из-за того, что в большинстве исследований приматов использовались электрофизиологические методы, в то время как при исследованиях у людей применялись функциональные методы нейровизуализации. Последний подход имеет низкое временное разрешение и усложняется присутствием сильного фонового «шума» сканера. Например, даже установление точных параметров человеческой тонотопической оси было проблемой, так как не-А1 слабо отвечает на чистые тоны, в то время как А1 отвечает кратковременно. Исследования, где была использована полосовая фильтрация стимулов со спектральными изменениями, убедительно показывают, что человеческая слуховая кора сохраняет тонотопическую ось. Корковая обработка височной информации зависит от скорости передачи информации. В общем, звуки могут быть подразделены на височные структуры замедленного темпа (<30 Гц) и ускоренного темпа (>50 Гц). По замедленному типу височная информация закодирована непосредственно невральными темпами выброса. И извилина Гешля, и не-А1 являются критическими элементами для модуляций замедленного уровня, неотъемлемой частью такого процесса как амплитудное детектирование речи. При ускоренном темпе, опережающем способности частоты невральной реакции, должны использоваться другие стратегии. Височные структуры высокой интенсивности отражаются в диапазоне миллисекунд, что играет роль для результатов исследований РПС (важный для локализации звука) и важно для восприятия музыкальных мелодий.
Несколько исследований показали, что мозг отвечает на височное вариации ускоренного темпа. Невральные реакции на постоянный шум выражены сильнее, чем на контрольный шум, так как высокочастотные колебания вызывают определенные реакции в А1. Только к середине XIX века удалось достигнуть согласия в том, что сенсорно-двигательные функции могут быть локализованы в коре головного мозга. Неприятие функциональной локализации отчасти было связано с реакцией на вводящие в заблуждение теории френологов, которые ограничивали связь сенсорно-двигательных функций только формой головы. В 1861 году при аутопсии пациента с экспрессивной афазией было обнаружено, что левая нижняя лобная извилина была поражена в области, которую с тех пор называют полем Брока. Повреждение задней верхней височной извилины в области, обычно называемой областью Вернике, приводит к сенсорному дефициту, который сразу проявляется клинически. Понятие тонотопии было определено в середине 1900-х, а многие предшествующие исследования специфической роли слуховой коры для частотной избирательности были неубедительны. В 1975 году было обнаружено, что двусторонние слуховые поражения коры приводят к потере способности локализовать источник звука. При удалении у обезьян части коры головного мозга, было определено, что двустороннее удаление верхней височной доли приводит к немедленной невосприимчивости звуковой стимуляции с отсроченным восстановлением в течение нескольких месяцев, в то время как одностороннее удаление приводило к контралатеральной недостаточности. Как было описано ранее, специфические особенности слухового поведения и невральной организации препятствуют нашему пониманию слуховой коры. Критические вопросы, касающиеся и нормальных, и патологических механизмов слуха остаются нерешенными. Конечной целью нейробиологии является понимание того, как работает головной мозг. По отношению к слуховой системе важно понимание того, как центральная и периферические слуховые системы обеспечивают слуховое распознавание и точное восприятие слуховых стимулов. До некоторой степени современное понимание было сформировано подходами, разделяющими центральную нервную систему на меньшие подгруппы и исследующими анатомию и физиология каждой подгруппы отдельно. Следовательно, мы знаем намного больше о каждой отдельной подгруппе, чем об их взаимодействии друг с другом. Кроме того, относительно мало известно, как именно подгруппы слуховых систем реагируют с другими сенсорными системами, чтобы обеспечить слуховое восприятие, которое будет правильно интерпретировано только в рамках реального контекста. Безусловно, полезно подразделить исследование слуховой системы на составляющие части, такие как А1 и вторичная слуховая кора, но у этого подхода есть внутренние ограничения, которые никогда не смогут привести к созданию удовлетворительной модели прослушивания для всего мозга. Представление о специфической «локализации» слуховой функции в отдельной нейроанатомической области, как кратко упоминалось выше, скорее всего слишком упрощенное. Целесообразнее определить функцию каждой подгруппы по порядку, чтобы получить возможность для интерпретации их функционирования в органическом контексте. Поэтому необходимо использование широкого спектра методов и развитие таких стратегий топографии распределения биопотенциалов как функциональная магнитно-резонансная томография (фМРТ), что даст важную информацию о взаимодействии слуховых областей друг с другом, а, возможно, и важные сведения об участии неслуховых областей в поведенческом контексте слушания. Одним из перимуществ фМРТ является то, что информация из разнообразных областей мозга может быть накоплена одновременно, позволяя фиксировать активность в тех областях мозга, связь которых со слуховым восприятием не предполагалась. Важно признать, что многие классические «неслуховые» области могут играть решающую роль при восприятии или генерации сложных звуков.
– Также рекомендуем “Функция и физиология слуховой коры” |
Источник
Зоны мозга, расположенные в корковом слое, отвечают за разные функции организма и способности человека. Взаимодействие всех отделов обеспечивает высшую психическую деятельность, в том числе мыслительные процессы, память и сознание, а также сложную двигательную активность. Благодаря слаженной работе корковых отделов человек способен обучаться, у него формируются мысли и эмоции, вырабатывается манера поведения.
Определение
Существует карта мозга, которая составлена немецким неврологом К. Бродманом и представляет собой описание мозговых корковых зон человека с выделением особенностей клеточного строения. Согласно карте, существует 52 поля, которые отличаются нейрональной организацией и функциями. Участки делятся на виды: первичные и вторичные, которые получают импульсы, передаваемые таламусом, и третичные, взаимодействующие исключительно с двумя первыми видами полей. Их функции:
- Первичные. Анализ нервных сигналов определенной модальности.
- Вторичные. Обеспечивают взаимодействие анализаторных (первичных) участков.
- Третичные. Определяют высшую психическую деятельность (мыслительные процессы, речь, интеллектуальные способности).
Различают ассоциативные и проекционные отделы в корковом слое. Основная задача ассоциативных – обеспечение взаимодействия между отдельными частями коры. Проекционные поддерживают связь между участками коры и подкорковыми структурами.
Структура коркового слоя и функции отделов
Продольная борозда разделяет мозг на большие полушария, которые состоят из 6 функциональных зон коры:
- Лобная.
- Теменная.
- Височная.
- Затылочная.
- Островок. Находится в углублении в Сильвиевой борозде.
- Лимбическая. Располагается с краю каждого полушария относительно срединной плоскости.
Несмотря на относительное разделение функций отдельных зон коры головного мозга, каждый физиологический процесс, протекающий в организме, требует их тесного взаимодействия и предполагает функциональную интеграцию. К примеру, зрительный центр располагается в затылочной области, однако в комплексном восприятии и обработке зрительного стимула кроме затылочной доли участвуют лобные и височные отделы.
Латерализация (процесс координации определенных функций разными полушариями) функций мозга предполагает относительное их разделение между 2 полушариями. К примеру, двигательные, осязательные, зрительные стимулы, поступающие из левой половины тела, перенаправляются в правое полушарие и наоборот. Некоторые сложные задачи оба полушария выполняют совместно, но наибольшее количество функций разделено между ними.
К примеру, левое доминирует в формировании речи, правое руководит в процессе обеспечения ориентации в пространстве. Первичные зоны коркового слоя головного мозга бывают моторными и сенсорными, другие отделы называют ассоциативными, которые еще известны как унимодальные и гетеромодальные. Унимодальные отделы находятся рядом с соответствующей сенсорной областью.
Они более тонко и глубоко обрабатывают информацию, поступающую в сенсорную область. Гетеромодальные отделы получают конвергентные (обладающие схожими признаками) данные из множества сенсорных и двигательных отделов. Благодаря работе гетеромодальных отделов происходит сравнение вновь поступающей информации с данными, хранящимися в инстинктивной и приобретенной памяти.
Корковый слой мозга разделен на зоны, которые в зависимости от расположения отвечают за выполнение разных задач, что позволяет выявлять участок поражения нервной ткани на основании симптомов. К примеру, зона Брока, известная как двигательный речевой центр, находится в височной части коркового слоя. Повреждение этого участка мозга провоцирует развитие моторной афазии. Пациент понимает речь, но не способен произносить слова, разговаривать.
Моторные отделы
Моторные зоны, находящиеся в коре больших полушарий, ответственны за выполнение волевых движений всех частей туловища. Благодаря участию ассоциативных отделов в процессе организации движений, осуществляется сложная, тонкая двигательная активность.
Двигательная зона (содержит поля Бродмана 4,6 и 8) коры, покрывающей головной мозг, расположена в передней части черепной коробки, пролегает перед линией центральной борозды, разделяющей доли мозга – лобную и теменную.
Этот участок контролирует и руководит осуществлением произвольных движений. Сигналы, поступающие из этого отдела мозга, приводят к сокращению мышц, пролегающих в пальцах рук и речевом аппарате (губы, язык), что обуславливает выполнение тонких движений. Этот участок моторной области обеспечивает способности человека:
- Произносить слова, говорить.
- Писать буквы, цифры и знаки.
- Умение играть на музыкальных инструментах.
Тонкой моторикой управляет большая часть двигательной области. Меньший участок нервной ткани регулирует сокращение и активность мышц спины, брюшного пресса и нижних конечностей. Этот участок моторной зоны обеспечивает устойчивость позы и выполнение крупных движений.
Сенсорные отделы
Сенсорные зоны (содержат поля Бродмана 1-3, 5 и 7) располагаются в дальних отделах коры, покрывающей большие полушария, отграниченных от лобного участка центральной извилиной. Эта доля, называемая теменной, содержит участок коры, который получает информацию от кожных рецепторов. Этот участок мозговой ткани обрабатывает информацию, формирующуюся при контакте кожи с посторонними предметами, водой, воздухом.
Благодаря деятельности этого отдела человек чувствует тепло, холод, ощущение от прикосновения при тактильном контакте, различает фактуру (шероховатую, острую или гладкую) и температуру (холодную или горячую) поверхности. В затылочной области находятся зрительные зоны коры, куда идут сведения от глаз. Зрительные нервы раздваиваются у основания мозга.
Одно ответвление отходит к противоположному полушарию. Обработкой сигналов, поступающих от органов зрения, занимаются поля Бродмана под номером 17-19. В поле 17 завершается центральный путь – здесь происходит оценка наличия и интенсивности импульсов, проходящих по волокнам зрительного нерва. В поле 18 и 19 осуществляется анализ таких параметров изображения, как цветовой оттенок, размеры, форма.
Слуховая зона расположена в височной области коры, покрывающей головной мозг, анализирует слуховые сигналы разной степени сложности. Карта мозга отводит слуховому отделу поля 22, 41 и 42. Здесь происходит оценка таких характеристик звука, как тембр, сила, громкость звучания, высота.
Благодаря деятельности этого отдела человек понимает с какой стороны поступает звуковой сигнал, определяет расстояние до источника звука, дифференцирует речь. Обонятельная система, как проекция в коре головного мозга, находится в поле 34. Вкусовой отдел занимает поле 43.
Ассоциативные зоны
Ассоциативные зоны находятся в коре, покрывающей большие полушария, не связаны с двигательными процессами или сенсорной деятельностью. Эти зоны головы занимают около 80% площади коры, преимущественно локализуются в долях – лобной и височной, а также затылочной и теменной. Каждый ассоциативный участок тесно взаимодействует с проекционными зонами, в том числе сенсорными и моторными, пролегающими в коре больших полушарий, образующих головной мозг.
Считается, что в этих отделах происходит объединение разрозненных фрагментов информации, в результате чего образуются сложные формы сознания. Проекционный участок опоясан ассоциативными областями, что обеспечивает взаимодействие, которое осуществляется посредством нейронов полисенсорной природы. Нервные клетки воспринимают сигналы, поступающие от разных органов и систем.
Они реагируют на информацию, передаваемую органами зрения и слуха, а также кожными рецепторами. Способность воспринимать информацию разного плана позволяет интегрировать данные, объединять их в общую систему, координировать двигательную и сенсорную деятельность. С ассоциативными зонами головного мозга и их функциями удобнее знакомиться при помощи таблицы.
| Отделы | Функции | Последствия поражения |
| Лобный | Высшая психическая деятельность – личностные характеристики, творчество, влечения | Утрата способности планировать и предвидеть, нарушение продуманного, целенаправленного поведения |
| Теменной | Формирование субъективной оценки окружающего пространства, получение представления о положении собственного тела и его перемещении | Утрата способности узнавать знакомые предметы при сохранении функциональности органов зрения |
| Височный | Речевая функция, распознавание и хранение речевой информации, воспроизводимой человеком и услышанной им | Утрата способности понимать чужую речь, невозможность распознавать слова при сохранении функциональности органов слуха |
Ассоциативные отделы, находящиеся в теменной доле, объединяют информацию, которая приходит от соматосенсорной системы. Соматосенсорная система образована рецепторами, обеспечивающими чувствительность, и центрами обработки информации, поддерживает сенсорные модальности, как температура, осязание, проприоцепция (ощущение своего тела – его положения, передвижения его частей), ноцицепция (физиологическая боль).
Ассоциативные отделы, находящиеся в височной части, отвечают за распознавание мелодий и разных сочетаний музыкальных звуков. Поле 37 позволяет запоминать слова. В височной части также содержатся центры сна, сновидений и памяти. Поле 39, расположенное на границе, разделяющей доли – височную, теменную, затылочную, содержит центр чтения, благодаря которому человек понимает письменный текст.
Патологии и признаки поражения отделов
Обширное поражение медиальных участков лобной доли провоцирует развитие абулии, которая проявляется замедленными реакциями, равнодушием, безучастностью к происходящему. При повреждении участка префронтальной орбитальной коры у пациента наблюдается отсутствие критической оценки собственного поведения, эмоциональная лабильность.
Двусторонняя травма в лобной области сопровождается признаками: ажитация, беспокойное поведение, навязчивость, многословие. Аномальное поведение является признаком деменции, которая развивается на фоне дегенеративных процессов, затрагивающих лобные доли. Повреждение мозгового вещества двигательной коры вызывает гемипарез или мышечную слабость.
Нарушения развиваются на стороне, противоположной местоположению патологического очага в мозге. Повреждение зрительного участка в одном полушарии приводит к развитию двухсторонней слепоты в половине поля обзора. Поражение поля 19 ассоциируется со зрительной агнозией – нарушением зрительного восприятия. Пациент видит предмет, но не может его распознать.
Информация, которая поступает через зрительный анализатор, не обрабатывается или обрабатывается неправильно, что приводит к невозможности различать знакомые предметы, лица людей. У таких больных нарушается цветовое восприятие – они не различают оттенки.
Повреждение поля 22 приводит к развитию музыкальной глухоты (нарушение восприятия музыкальных произведений), появлению слуховых галлюцинаций, нарушению реакций, ориентированных на слуховые раздражители. Поражение поля 41 сопровождается развитием корковой глухоты (невозможность восприятия звуковых сигналов).
Поражение поля 34 сопровождается нарушением восприятия запахов, в том числе обонятельными галлюцинациями. Патологические структурные изменения в нервной ткани поля 39 приводят к неспособности читать и писать. При повреждении ткани поля 37 человек не помнит названия предметов.
Зоны мозга разделяются на сенсорные и двигательные, а также ассоциативные – причем все участки взаимодействуют между собой. Каждый отдел наделен определенными функциями, которые в совокупности обуславливают высшую психическую и сложную двигательную деятельность.
Просмотров: 448
Источник