Онтогенез кори головного мозку

В развитии центрального отдела нервной системы имеют большое значение 3 процесса:
1. пролиферация
2. миграция
3. дифференцировка
Пролиферация начинается с 18-х суток эмбриогенеза, при этом в свернувшейся нервной трубке выделяют один слой, содержащий два вида стволовых клеток для образования двух основных дифферонов нервной ткани (первый – для развития нейронов, второй – для развития макроглиоцитов). Стволовые клетки разделяют между собой особые клетки – эмбриональные радиальные эпендимоциты или танициты. Танициты создают внутренние и наружные разграничительные мембраны. Пролиферация (размножение нервных клеток) завершается во второй половине беременности, но уже к 21 суткам в составе нервной трубки выделяют 3 слоя клеток:
1. Вентрикулярный (внутренний)
2. Субвентрикулярный
3. Маргинальный
В первом и во втором слое содержатся стволовые и полустволовые клетки обоих дифферонов, которые размножаются митотически с одинаковой скоростью (20 тысяч клеток в минуту), что приводит к образованию 150 млрд. нейронов только для будущей коры головного мозга и столько же глиальных клеток. Количество делений у пролиферирующих клеток запрограммировано и затем они начинают активно мигрировать для формирования маргинального слоя.
Миграция происходит под регуляцией таницитов и только по их поверхности благодаря выделению ими факторов миграции и гликонектина. Нейроны начинают ползти вверх по отростку таницита в маргинальный слой и остаются там, на определенном месте, которое также регулирует таницит, при этом, по мере продвижения клеток, может быть остановка и группировка нейронов с формированием мозговых ядер. Вторая разновидность миграции в конечном мозге создает скопление нейронов под наружной глиальной мембраной в виде кортикальной пластинки. При формировании кортикальной пластинки все пришедшие нейроны в ее состав начинают дифференцироваться, то есть усложняют строение тела нейрона и его отростков. Скорость роста аксона очень большая и достигает 1-2 мм в час. Пришедшие в кортикальный слой нейроны располагаются строго упорядочено между двумя отростками соседних таницитов в виде цепочек или столбиков. Таким создавшимся цепочкам нейронов дали название онтогенетическая гистологическая колонка. Однотипные после дифференцировки нейроны в колонках в результате миграции останавливаются на одном и том же уровне, создавая эффект послойного расположения нейронов в коре полушарий. Все нейроны обязательно вступают друг с другом во взаимосвязь, те нейроны, которые не сформировали синапсы, а это 87-90% клеток сразу же уничтожаются особым фактором мужания, вырабатываемым таницитами. Количество нейронов, формирующих кору, в конечном счете, это 15-10 млрд. Эмбриональные танициты после выполнения своей функции также уничтожаются.
Цитоархитектоника, миелоархитектоника и модульный принцип организации коры головного мозга
В головном мозге различают серое и белое вещество, но их распределение здесь значительно сложнее, чем в спинном мозге. Большая часть серого вещества головного мозга располагается на поверхности большого мозга и мозжечка, образуя их кору толщиной 3-5 мм. Меньшая часть образует многочисленные подкорковые ядра, окруженные белым веществом. Все серое вещество состоит из мультиполярных нейронов.
Цитоархитектоника
Нейроны коры расположены нерезко отграниченными слоями, которые обозначаются римскими цифрами и нумеруются снаружи внутрь. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В коре полушарий различают шесть основных слоев:
· I – молекулярный;
· II – наружный зернистый;
· III – пирамидный;
· IV – внутренний зернистый;
· V – ганглионарный;
· VI – слой полиморфных клеток.
I – Молекулярный слой коры содержит множество отростков и небольшое количество мелких ассоциативных горизонтальных клеток Кахаля, нейроны с аксональной кисточкой (по функции тормозные). Их аксоны проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Однако основная масса волокон этого сплетения представлена ветвлениями дендритов нижележащих слоев.
II – Наружный зернистый слой образован многочисленными мелкими пирамидными и звездчатыми шипиковыми нейронами (по функции возбуждающими), а также тормозными нейронами, к которым относятся малые и большие корзинчатые клетки, нейроны с аксоаксональными синапсами. Дендриты этих клеток поднимаются в молекулярный слой, а аксоны либо уходят в белое вещество, либо, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя.
III – Самый широкий слой коры большого мозга – пирамидный. Он содержит пирамидные нейроны, клетки Мартинотти и клетки с двойным букетом дендритов (они являютя тормозными для тормозных нейронов). Апикальные дендриты пирамид уходят в молекулярный слой, боковые дендриты образуют синапсы со смежными клетками этого слоя. Аксон пирамидной клетки всегда отходит от ее основания. У мелких клеток аксон остается в пределах коры, у крупных он формирует миелиновое волокно, идущее в белое вещество головного мозга. Пирамидный слой выполняет преимущественно ассоциативные функции. Аксоны пирамидных нейронов этого слоя образуют кортико-кортикальные пути.
IV – Внутренний зернистый слой в некоторых полях коры развит очень сильно (например, в зрительной и слуховой зонах коры), а в других он может почти отсутствовать (например, в прецентральной извилине). Этот слой образован мелкими звездчатыми шипиковыми нейронами двух типов: фокального и диффузного. В его состав входит большое количество горизонтальных волокон.
V – Ганглионарный слой коры образован крупными пирамидами, причем область моторной коры (прецентральная извилина) содержит гигантские пирамиды, которые впервые описал киевский анатом В. Бец. Апикальные дендриты пирамид достигают I-го слоя. Аксоны пирамид проецируются на моторные ядра головного и спинного мозга. Наиболее длинные аксоны клеток Беца в составе пирамидных путей достигают каудальных сегментов спинного мозга. Кроме пирамидных нейронов в ганглионарном слое коры встречаются вертикальные веретеновидные клетки, а также малые и большие корзинчатые клетки.
VI – Слой полиморфных клеток образован разнообразными по форме нейронами (веретеновидными, звездчатыми, клетками Мартинотти). Аксоны этих клеток уходят в белое вещество в составе эфферентных путей, а дендриты достигают молекулярного слоя.
Миелоархитектоника
Среди нервных волокон коры полушарий большого мозга можно выделить ассоциативные волокна, связывающие отдельные участки коры одного полушария, комиссуральные, соединяющие кору различных полушарий, и проекционные волокна, как афферентные, так и эфферентные, которые связывают кору с ядрами низших отделов центральной нервной системы.
В коре полушарий проекционные волокна образуют радиальные лучи, заканчивающиеся в III – пирамидном слое. Кроме уже описанного тангенциального сплетения I – молекулярного слоя, на уровне IV – внутреннего зернистого и V – ганглионарного слоев расположены два тангенциальных слоя миелиновых нервных волокон – соответственно, наружная полоска Байярже и внутренняя полоска Байярже.
Источник
Уже в начале 4-го месяца у зародыша человека большие полушария покрывают зрительные бугры. Их поверхность в это время еще гладкая. На ней есть только вдавление — будущая сильвиева борозда. К 5-м годам борозды еще не достигают полной глубины и имеются отличия в расположении долей мозга. Приблизительно к 9-10 годам борозды и извилины располагаются так же, как у взрослого.
В онтогенезе сначала появляются глубокие слои коры, а затем уже поверхностные. Области, различные но филогенетическому происхождению, в онтогенезе развиваются различно. Уже на ранних ступенях внутриутробного развития большие полушария превосходят по величине остальные отделы мозга. К концу 7-го месяца они покрывают весь мозжечок и в развитом мозге взрослого человека составляют более 80% веса головного мозга. Развитие нейронов коры предшествует в онтогенезе развитию борозд. Предполагается, что у новорожденного ребенка почти такое же количество нейронов в головном мозге, как у взрослого, так как после рождения, по-видимому, появляется мало нейронов в отличие от размножения клеток нейроглии. После рождения, особенно до 3 лет, нейроны дифференцируются, увеличивается число дендритов и их разветвлений, шипиков и синапсов. При определенных условиях растут ответвления и от аксонов; последние, как и дендриты. покрываются миелином. В коре головного мозга большая часть пресинаптических волокон не миелинизирована, только 1/8 их часть миелинизирована, что обеспечивает быстрое проведение импульсов возбуждения. С возрастом относительный объем тел нейронов уменьшается, а объем дендритов увеличивается и возрастает их поверхность, тогда как поверхность тел нейронов остается почти постоянной, начиная с грудного возраста. На 6-м месяце утробной жизни 95-96% коры больших полушарий состоит из 6 слоев. У ребенка 3 лет нейроны коры отчетливо дифференцированы и к 8 годам мало отличаются от нейронов взрослого. Рост пирамидных нейронов лобных долей больших полушарий происходит у детей неравномерно. Приблизительно к 12 годам он заканчивается в нижних слоях коры, а в верхних ее слоях некоторое ускорение роста пирамидных нейронов продолжается до 18 лет.
С возрастом дифференцируется и подкорковый отдел двигательного анализатора. Примерно к 7 годам кинестезический анализатор имеет такое же строение, как у взрослого, но он продолжает развиваться. Зрительный анализатор формируется несколько позже двигательного и кинестезического. Он дифференцируется к 7-12 годам, но его развитие также продолжается. Строение нейронов коры и их отростков усложняется примерно до 40 лет и позже.
У новорожденных масса головного мозга приближается к 400 г. В 2 месяца ребенок реагирует голосом на обращение взрослых и масса мозга увеличивается до 480 г. В 6 месяцев он подражает звукам и масса мозга доходит до 660 г. В 9-11 месяцев, когда ребенок начинает говорить, масса мозга повышается до 770-850 г. В связи с быстрым увеличением словарного запаса к 13-17-21 месяцам масса мозга уже равна 930-1030-1060 г. Следовательно, развитие головного мозга детей, особенно его рост, резко ускоряется в связи с формированием речи и умственной деятельности, оно взаимосвязано с усложнением моторики и психики. Развитие головного мозга, в особенности дифференцирование и рост нейронов, в младшем школьном возрасте происходит быстрее, чем в последующие годы. До 2 лет нет различия в функциях обоих больших полушарий, а затем оно начинает проявляться. У трети людей полушария не приобретают четкой функциональной специализации, которая формируется только в процессе обучения и воспитания.
Источник
Эмбриогенез головного мозга (этапы развития и формирования)
а) Эмбриогенез структур мозга. В конце 4-й недели ростральная часть нервной трубки образует изгиб на уровне будущего среднего мозга. Эта область — средний мозговой пузырь — легкими перетяжками отграничена от переднего мозгового пузыря (будущий передний мозг) и ромбовидного мозгового пузыря (будущий задний мозг).
Крыловидная пластинка переднего мозгового пузыря расширяется и образует конечный мозг (будущие полушария головного мозга). Базальная пластинка сохраняет свое положение и дает начало промежуточному мозговому пузырю. В промежуточном мозговом пузыре формируются глазные ямки — закладки сетчатки и зрительного нерва.
Промежуточный, средний и ромбовидный мозговые пузыри формируют зачаточный ствол головного мозга.
По мере развития ствол мозга изгибается, в результате чего средний мозговой пузырь перемещается наверх. Ромбовидный мозговой пузырь образует складку так, что крыловидная пластинка расходится и формирует четвертый желудочек мозга ромбовидной формы. Ростральная часть ромбовидного мозгового пузыря дает начало мосту мозга и мозжечку, а каудальная — продолговатому мозгу.
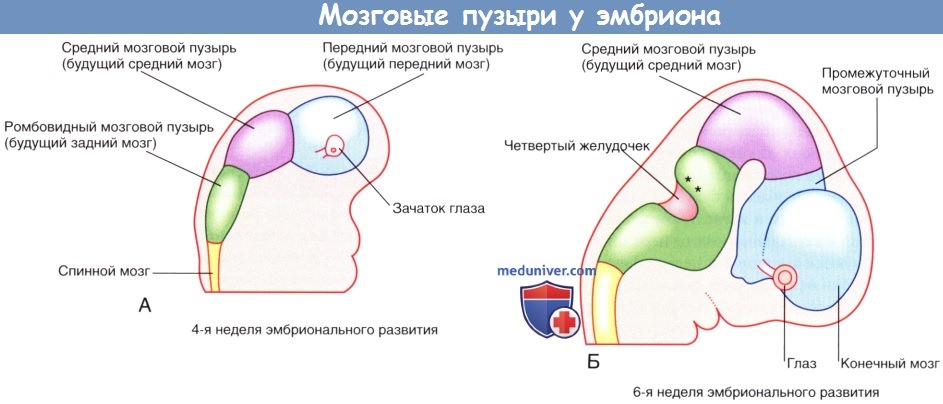
(А и Б) Мозговые пузыри, вид справа. Звездочками обозначены участки первичного развития мозжечка.
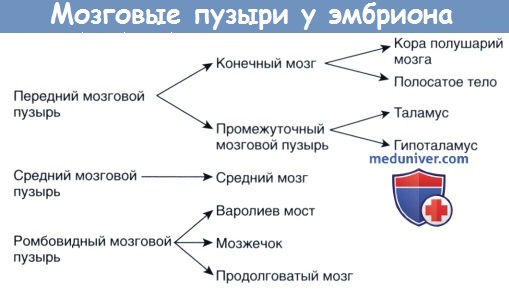
Некоторые производные мозговых пузырей.
б) Эмбриогенез системы желудочков мозга и сосудистого сплетения. Канал нервной трубки расширяется и образует полушария мозга, в которых формируются боковые желудочки. Боковые желудочки сообщаются с третьим желудочком, структуры которого окружает промежуточный мозговой пузырь. Третий желудочек сообщается с двумя боковыми желудочками через отверстие Монро (межжелудочковое отверстие). Третий и четвертый желудочки сообщаются через водопровод мозга (сильвиев водопровод), расположенный внутри среднего мозга.
Пучки капилляров инвагинируют тонкие верхние стенки переднего и заднего мозга и образуют сосудистое сплетение четырех желудочков мозга. Клетки сосудистого сплетения секретируют спинномозговую жидкость, которая поступает в систему желудочков и затем оттекает из IV желудочка через три отверстия в его крыше.
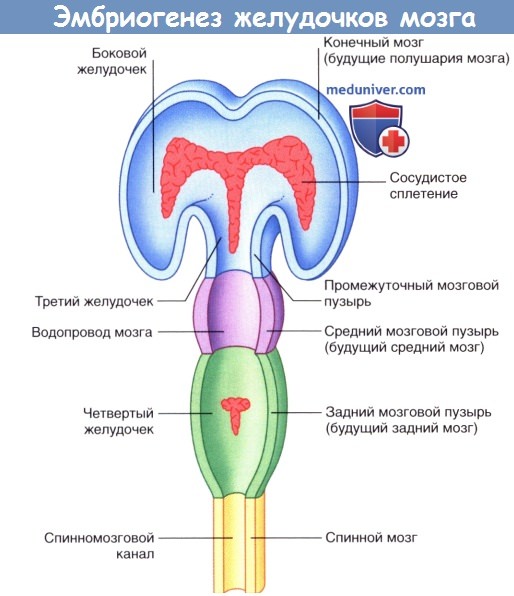
Развитие системы желудочков мозга. Красным цветом показаны сосудистые сплетения.
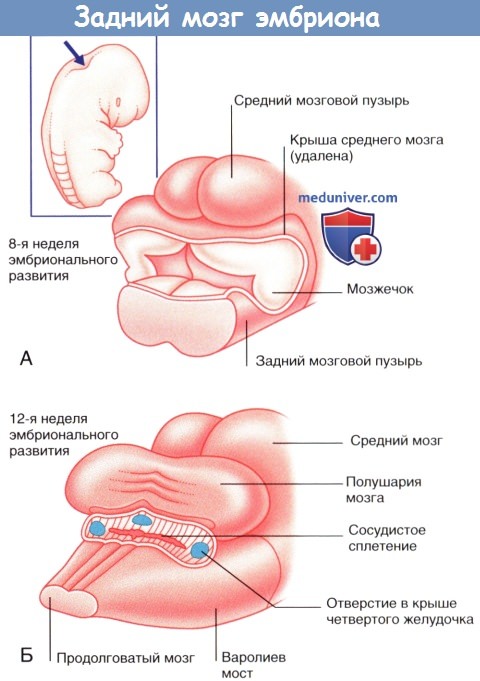
Вид с дорсальной стороны на развивающийся задний мозг (указано стрелкой).
(А) На 8-й неделе развития четвертый желудочек дает начало мозжечку.
(Б) На 12-й неделе мозжечок скрывает четвертый желудочек, и на его крыше появляются три отверстия.
в) Эмбриогенез черепных нервов. На рисунке ниже показана стадия развития черепных нервов на 6-й неделе после оплодотворения.
• Обонятельный нерв (I) образуется из биполярных нейронов, развивающихся из эпителиальных клеток обонятельной ямки.
• Зрительный нерв (II) формируется из сетчатки.
• Глазодвигательный (III) и блоковый (IV) нервы развиваются из клеток среднего мозга, а отводящий (VI) —из клеток варолиевого моста.
• II, III и IV нервы в дальнейшем будут иннервировать наружные мышцы глаза.
• Три ветви тройничного (V) нерва будут обеспечивать чувствительность кожи лица и головы, полостей рта и носа, а также зубов. Двигательные ветви предназначены для иннервации жевательных мышц.
• Лицевой (VII) нерв будет обеспечивать иннервацию мимических мышц лица.
• Преддверно-улитковый (VIII) нерв будет отвечать за передачу нервных импульсов к органам слуха и равновесия, образующихся из слухового пузырька.
• Языкоглоточный (IX) нерв — смешанный. Большая часть его волокон обеспечивает чувствительность ротоглотки и двигательную иннервацию шилоглоточной мышцы.
• Блуждающий нерв (X) — также смешанный, обеспечивает обильную чувствительную иннервацию слизистой оболочки пищеварительного тракта и значительную двигательную (парасимпатическую) иннервацию сердца, легких и желудочно-кишечного тракта.
• Церебральная часть добавочного нерва (ХIс) в составе блуждающего нерва будет подходить к мышцам гортани и глотки.
• Спинальная часть добавочного нерва (XIs) будет иннервировать грудино-ключично-сосцевидную и трапециевидную мышцы.
• Подъязычный нерв (XII) будет иннервировать все мышцы языка, за исключением небно-язычной, движения которой обеспечивают нервы глоточного сплетения.
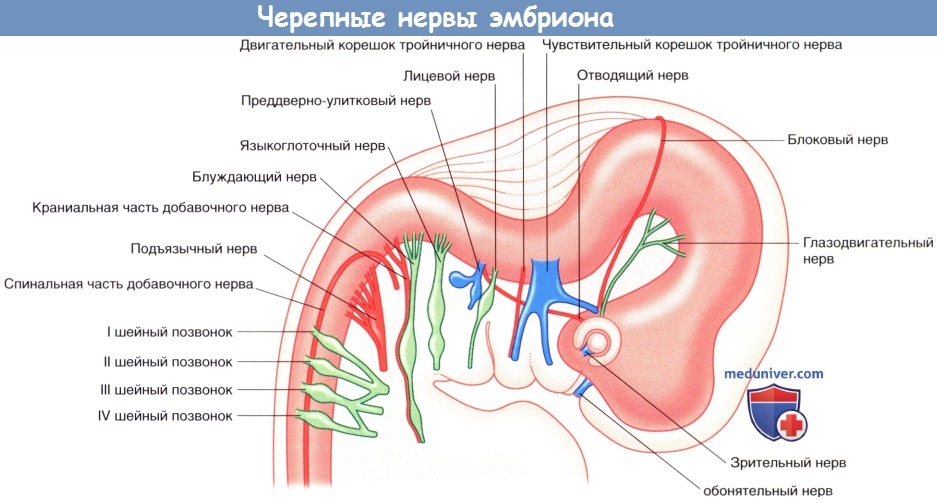
Черепные нервы шестинедельного эмбриона.
г) Эмбриогенез полушарий мозга. В желудочковой зоне конечного мозга, снаружи от бокового желудочка, клетки делятся и мигрируют на внешнюю поверхность развивающихся полушарий, формируя корковое вещество полушарий мозга.
Полушария мозга увеличиваются неравномерно. Относительно «неактивным» остается островок мозга. Эта структура образует опорную точку, вокруг которой происходит ротация полушарий. Лобную, теменную, затылочную и височные доли можно различить на 14-й неделе гестации.
На медиальной поверхности полушарий расположен гиппокамп — участок коры головного мозга, который относят к пятой доли мозга — лимбической. Гиппокамп перемещается в височную долю, оставляя за собой след, представленный нервными тяжами, получившими название свод мозга. Под этой дугой располагается сосудистая щель, через которую сосудистое сплетение погружается в боковой желудочек.
Передняя комиссура соединяет обонятельные зоны слева и справа. Важно отметить, что более крупная комиссура — мозолистое тело — соединяет соответствующие друг другу зоны коры двух полушарий мозга. Мозолистое тело расширяется в заднем направлении над стволом мозга.
На коронарных срезах конечного мозга в основании каждого полушария можно увидеть скопления серого вещества, являющиеся предшественниками полосатого тела. Вблизи III желудочка промежуточный мозговой пузырь развивается в таламус и гипоталамус.
Увеличивающиеся в размере полушария мозга соприкасаются с промежуточным мозговым пузырем, и происходит их срастание (см. «участок срастания» на рисунке ниже). Вследствие этого, во-первых, выделяют понятие «ствол мозга», включающее структуры, оставшиеся свободными: средний мозг, мост и продолговатый мозг. Во-вторых, кора больших полушарий соединена нервными волокнами непосредственно со стволом мозга. Вместе с нервными волокнами, идущими от таламуса к коре, эти волокна разделяют полосатое тело на хвостатое ядро и чечевицеобразное ядро.
К 28-й неделе эмбрионального развития на поверхности мозга появляются некоторые борозды, в частности латеральная, центральная и шпорная.
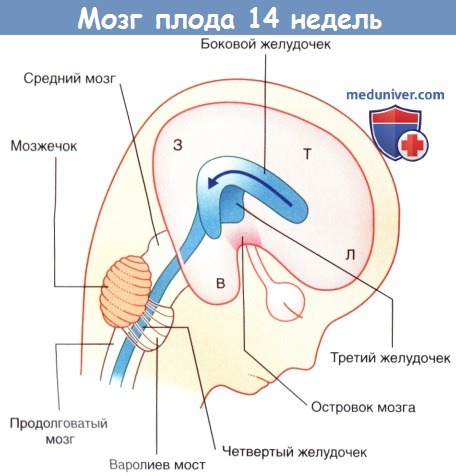
Мозг плода на 14-й неделе развития.
Стрелкой показан С-образный рост полушарий вокруг островка мозга.
Л—Лобная доля. Т—Теменная доля. З—Затылочная доля. В—Височная доля.
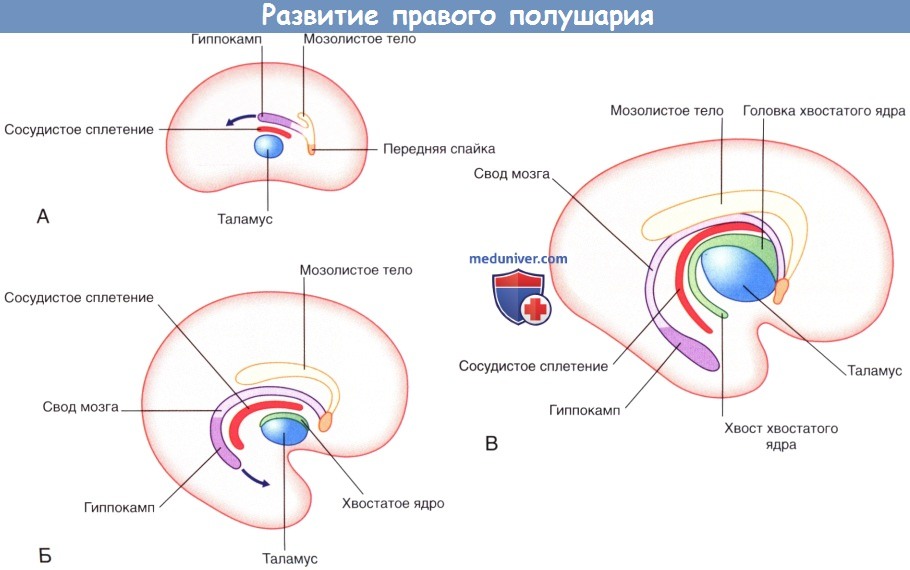
Развитие правого полушария (вид с медиальной стороны).
Гиппокамп, изначально расположенный дорсапьно по отношению к таламусу, мигрирует в височную долю (на рисунках А и Б обозначено стрелками), оставляя за собой след, — свод мозга.
Таким образом формируется дуга, под которой располагаются щель сосудистого сплетения (место его вхождения в желудочек) и хвост хвостатого ядра.
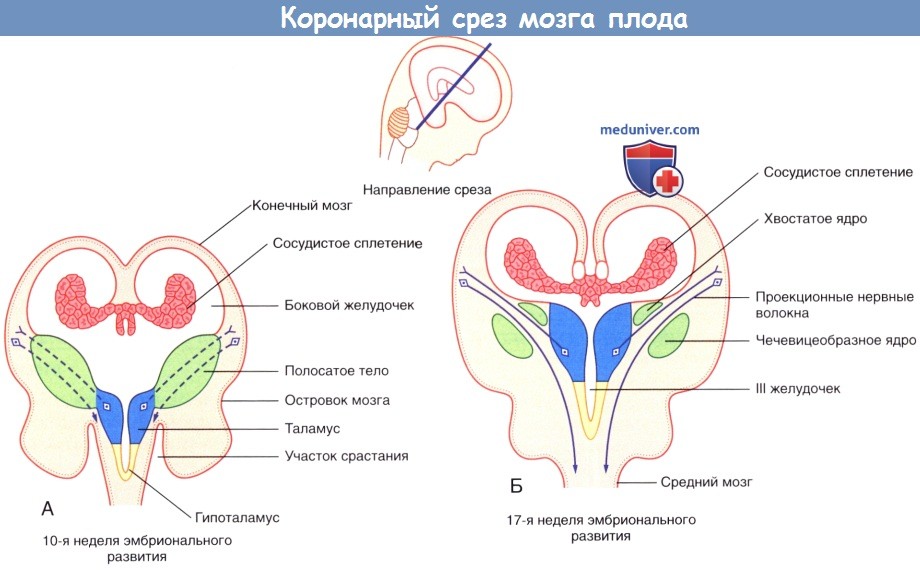
Коронарный срез мозга плода.
(А) Волокна, соединяющие таламус с корой и кору со спинным мозгом, пересекают полосатое тело на 10-й неделе развития эмбриона.
(Б) Полосатое тело разделяется на хвостатое и чечевицеобразное ядра на 17-й неделе развития плода.
Слияние сохраняется на переднем конце (на рисунке не показано).
Три основные борозды у 28-недельного плода.
(А) Латеральная и (Б) медиальная поверхности левого полушария мозга.
д) Резюме. Нервная система формируется из нервной трубки эктодермального происхождения, полостью которой служит спинномозговой канал. Некоторые клетки мигрируют по разные стороны от трубки и формируют нервный гребень. Часть нервной трубки, расположенная наиболее каудально, дает начало спинному мозгу. Из нервного гребня образуются клетки спинального ганглия, от которого корешки спинномозговых нервов отходят к чувствительной крыловидной пластинке. Базальная мозговая пластинка представлена двигательными нейронами, от которых отходят передние корешки. Объединяясь с задними корешками, передние корешки образуют спинномозговые нервы.
Наиболее рострально расположенная часть нервной трубки образует три желудочка мозга. Передний мозговой пузырь (будущий передний мозг) в дорсальной части дает начало полушариям мозга (конечный мозг), а в вентральной — промежуточному мозговому пузырю. Средний мозговой пузырь формирует средний мозг. Ромбовидный мозговой пузырь образует задний мозг, представленный мостом, продолговатым мозгом и мозжечком.
Нервная трубка расширяется в ростральном направлении и формирует систему желудочков мозга. Спинномозговую жидкость секретируют клетки сосудистого сплетения, инвагинирующие верхние стенки желудочков.
Полушария мозга формируют лобную, теменную, височную, затылочную и лимбинескую доли. Полушария мозга образуют поперечные связи с мозолистым телом, а также с передними и задними комиссурами. Скопления серого вещества, располагающиеся в основании каждого полушария, дают начало полосатому телу. Полушария мозга срастаются с боковыми стенками промежуточного мозгового пузыря, после чего средний и задний мозговые пузыри становятся единственными остатками ЦНС, характерной для эмбриона.
– Также рекомендуем “Топография и характеристика поверхности полушарий мозга”
Редактор: Искандер Милевски. Дата публикации: 9.11.2018
Источник