Анатомія кори головного мозку

Окремі ділянки кори мають різне функціональне значення. Разом з підкірковими центрами, стовбуром мозку і спинним мозком великий мозок об’єднує окремі частини організму в єдине ціле, здійснює нервову регуляцію всіх органів (рис. 55, 56).
У кору великого мозку надходять доцентрові імпульси від рецепторів. Кожному рецепторному апарату відповідає в корі ділянка, яку І.П Павлов назвав кірковим ядром аналізатора. Ділянка кори, де розташовані кіркові ядра аналізаторів, названі сенсорними зонами кори великого мозку.
Ядерна зона рухового аналізатора (сомато-сенсорна зона), куди надходять збудження від рецепторів суглобів, скелетних м’язів і сухожилок, розташована в передню- і задньоцентральних ділянках кори. У межах передньої центральної закрутки найвище розміщені центри для м’язів нижньої кінцівки, нижче – для м’язів тулуба, потім верхньої кінцівки і, нарешті, центри м’язів голови. Ураження цієї зони призводить до паралічу протилежної половини тіла.
Рис. 55. Зовнішня будова головного мозку (за дорлінг кіндерслі, 2003)
Передцентральна звивина (gyrus precentralis) і прицентральна часіка (lobulus paracentralis) лобової частки становлять руховий центр кори і є аналізатором кінестезичних імпульсів, які надходять від посмугованих м’язів, суглобів, сухожилків. Тут замикаються рухові умовні рефлекси. У верхній ділянці перед центральної звивини розташовані клітинні групи, що належать до м’язів нижніх кінцівок, нижче – верхніх кінцівок, ще нижче – неврони, пов’язані з іннервацією м’язів голови. Оскільки нервові шляхи перехрещуються, праві рухові центри кори пов’язані з мускулатурою лівої сторони тіла і навпаки.
У задній частині середньої лобової звивини міститься центр узгодженого руху голови й очей (окоруховий, блоковий, відвідний і додатковий нерви).
Рис. 56. Кіркові поля (за Дорлінг Кіндерслі, 2003)
У задньому відділі нижньої лобової звивини розміщена зона Брока – руховий центр мови, який разом із центром Верніке забезпечують здатність людини читати, писати, чути, вимовляти і розуміти мову.
Ушкодження різних полів кори лобової частки може призвести: до підвищення агресивності й послаблення реакції страху; зростання пасивно-захисних умовних рефлексів; порушення харчових і захисних умовних рефлексів.
У таких людей спостерігається втрата ініціативи, апатія, порушення абстрактного мислення, нездатність до творчого мислення, розгальмування нижчих емоцій і потягів, розлади мовлення і понятійного мислення.
У задній частині лобової звивини розташований центр письма, ураження якого призводить до порушення навичок письма під контролем зору.
У лівій (у лівшів у правій) нижній тім’яній часточці розташований центр, який координує цілеспрямовані рухи. Він функціонує за типом тимчасових зв’язків, які виникають протягом індивідуального життя, тобто умовних рефлексів. У разі ушкодження цього центру елементи довільних рухів зберігаються, але порушуються цілеспрямовані дії (апраксія).
У верхній тім’яній частці (задньоцентральна звивина) розміщений кірковий центр аналізаторів чутливості (больової, температурної, дотикової), або сомато-сенсорна кора. Ураження кори у цій частині призводить до часткової або повної анестезії (втрата чутливості).
Ураження кори в ділянці верхньої тім’яної частки призводить до зниження больової чутливості і порушення стереогноза – впізнавання предметів на дотик без допомоги зору.
У нижній тім’яній частині розташований центр праксії, який регулює здатність здійснювати координаційні рухи, які складають основу робочих процесів і потребують спеціального навчання.
У кутовій звивині тім’яної частки розташований зоровий центр мови. Його ураження призводить до неможливості розуміння письма (алексія).
Тім’яна ділянка – це апарат вищої інтегративної діяльності мозку людини, вона безпосередньо стосується процесів біологічної і соціальної адаптації, є фізіологічною основою вищих психічних функцій.
Локалізація статичного аналізатора (центр збереження рівноваги і положення тіла в просторі) – кора верхньої та середньої скроневих звивин. Ушкодження цього центру призводить до атаксії (розладу координації рухів).
Зона шкірного аналізатора, зв’язаного з температурою, больовою і тактильною чутливістю займає задньоцентральну ділянку. Центри чутливості нижчих частин тіла розміщені у верхніх частинах тіла – у нижніх її ділянках.
Найбільшу площу займає кіркове представництво рецепторів кисті рук, голосового аналізатора і обличчя, найменшу – тулуба, стегна і гомілки.
Ядерна зона зорового аналізатора розташована на внутрішній поверхні потиличної ділянки, в зоні шпорної борозни. Ураження цього центру призводить до сліпоти. При порушеннях у сусідніх із шпорною борозною частин кори в ділянці потиличного полюса на медіальній і латеральній поверхнях частки може спостерігатися втрата зорової пам’яті, здатності орієнтації у незнайомій обстановці, порушення функції, пов’язаної із бінокулярним зором (здатності за допомогою зору оцінювати форму предметів, відстань до них тощо).
У корі верхньої скроневої звивини розташована частина слухового аналізатора, а поблизу від бокової борозни – ядерна зона смакового аналізатора. Двостороннє ураження до повної кіркової глухоти.
Нюхова зона розміщена на внутрішній поверхні скроневих ділянок кори. В ділянці середньої і нижньої скроневих звивин розташоване кіркове представництво вестибулярного аналізатора. Ураження цієї ділянки призводить до порушення рівноваги під час стояння і зниження чутливості.
Із сенсорними зонами взаємодіє моторна зона кори великого мозку. Ядерні зони аналізаторів – це ділянки кори, в яких закінчується основна маса провідних шляхів аналізаторів. За межами ядерних зон розташовані розсіяні елементи, куди надходять імпульси від тих же рецепторів, що і в ядро аналізатора.
Центр мови міститься у лівій півкулі. Розрізняють 2 центри мови: руховий (зона Брока), який міститься у нижній частині лобової ділянки і слуховий (зона Верніке), який знаходиться у скроневій ділянці, під заднім кінцем сільвієвої борозни. Центри мови є лише у людини. Мовлення, мислення, почуття і вправні рухи контролюються нейронами, які розміщені в лобовій ділянці головного мозку. Розпізнавання тонів і звуків відбувається в скроневій ділянці. Ця ділянка також бере участь у запам’ятовуванні інформації. Різноманітні сенсорні відчуття, такі як біль, температура усвідомлюються та інтерпретуються в тім’яній ділянці. Потилична ділянка фіксує та інтерпретує зорові образи.
Источник
Кора головного мозга. Физиологическая анатомия коры головного мозга
По иронии судьбы, из всех отделов головного мозга меньше всего мы знаем о функциях коры большого мозга, хотя совершенно очевидно, что это самая большая часть нервной системы. В тоже время известны эффекты поражения или специфической стимуляции различных участков коры. В первых статьях по физиологии на сайте обсуждаются фактические данные о функциях коры; затем кратко представлены основные теории о нервных механизмах, участвующих в мыслительных процессах, памяти, анализе сенсорной информации и др.
а) Физиологическая анатомия коры большого мозга. Функциональном частью коры большого мозга является гонкий слои нейронов, покрывающий поверхность всех извилин. Толщина этого слоя составляет лишь 2-5 мм, а общая площадь равна примерно 0,25 м. В целом кора большого мозга содержит примерно 100 млрд нейронов.
На рисунке ниже показана типичная гистологическая структура коры большого мозга с последовательными слоями разных типов нейронов.
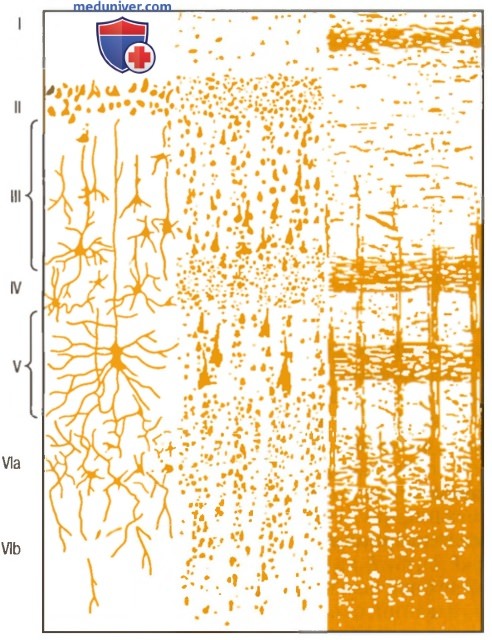
Структура коры большого мозга.
I – молекулярный слой; II — наружный зернистый слой;
III – слой пирамидных клеток; IV – внутренний зернистый слой;
V – слой больших пирамидных клеток. VI – слой веретенообразных клеток
Большинство нейронов относят к трем типам:
(1) зернистые (также называемые звездчатыми);
(2) веретенообразные,
(3) пирамидные, получившие свое название за характерную пирамидальную форму.
Зернистые нейроны обычно имеют короткие аксоны и, следовательно, функционируют в основном как вставочные нейроны, которые проводят нервные сигналы только на небольшие расстояния в пределах самой коры. Некоторые нейроны — возбуждающие, они выделяют в основном нейромедиатор глутамат; другие — тормозящие, основным непромедиатором которых является гамма-аминомасляная кислота — главный тормозящий нейромедиатор.
Сенсорные области коры, а также ассоциативные, расположенные между сенсорными и моторными областями, содержат большое количество зернистых клеток, что дает основание предполагать высокую степень внутрикоргикалыюй обработки входящих сенсорных сигналов в пределах сенсорных и ассоциативных областеи.
Пирамидные и веретенообразные клетки дают начало почти всем волокнам, выходящим из коры. Пирамидные клетки — более крупные и мно гочисленпые, чем веретенообразные. Они являются источником длинных толстых нервных волокон, которые идут, не прерываясь, в спинной мозг. Они также дают начало крупным подкорковым пучкам ассоциативных волокон, проходящим от одной крупной части головного мозга к другой.
Справа на рисунке выше показана типичная организация нервных волокон в пределах разных слоев коры большого мозга. Обратите особое внимание на большое число горизонтальных волокон, которые распространяются между при лежащими областями коры. Также отметьте вертикальные волокна, идущие к коре и от нее к нижележащим областям мозга. Некоторые из них проходят весь путь до спинного мозга или направляются к отдаленным регионам коры большого мозга через длинные ассоциативные пучки.
Функции специфических слоев коры большого мозга обсуждаются в отдельных статьях на сайте (просим вас пользоваться формой поиска выше). Здесь следует вспомнить, что большинство входящих специфических сенсорных сигналов от тела поступают в IV слой коры. Большая часть сигналов покидают кору через нейроны, локализованные в V и в VI слоях; очень крупные волокна, идущие к мозговому стволу и спинному мозгу, берут начало главным образом в V слое, а громадное количество волокон, идущих к таламусу, возникают в VI слое.
Слои I, II и III осуществляют основную часть ассоциативных функций внутри коры, причем особенно большое число нейронов II и III слоев формируют короткие горизонтальные связи с прилежащими областями коры.
– Также рекомендуем “Связи коры головного мозга с другими отделами. Специфические области коры большого мозга”
Оглавление темы “Головной мозг и его физиология”:
1. Кора головного мозга. Физиологическая анатомия коры головного мозга
2. Связи коры головного мозга с другими отделами. Специфические области коры большого мозга
3. Ассоциативные области коры головного мозга. Физиологические отделы мозга
4. Префронтальная ассоциативная область. Лимбическая ассоциативная область
5. Область распознавания лиц. Височная доля и область Вернике
6. Концепция доминантного полушария. Роль языка в функции области Вернике
7. Функции теменно-затылочно-височной коры. Высшие интеллектуальные функции
8. Сниженная агрессивность. Развитие мысли
9. Функция мозга в коммуникации. Сенсорные и моторные аспекты коммуникации
10. Артикуляция. Функция мозолистого тела и передней спайки
Источник
Взаємовідносини борозен і звивини з кістками і швами черепа у новонародженої дитини інші, ніж у дорослого. Основні борозни (центральна, латеральна) виражені добре, але гілки основних борозен і дрібні звивини позначені слабо. Надалі в процесі розвитку кори борозни стають глибшими, а звивини між ними рельєфніше. Співвідношення борозен, звивин і швів черепа, характерне для дорослої людини, встановлюється у дітей в 6-8 років.
Протягом перших місяців життя розвиток кори йде дуже швидкими темпами. Більшість нейронів набувають зрілу форму, інтенсивно відбуваються процеси мієлінізації нервових волокон, що дозволяють реагувати на зовнішні подразники більш диференційовано.
У процесі еволюції людини як біологічного виду, а також у процесі онтогенезу – індивідуального розвитку кожної людини – відбувається кортікалізаціі функцій, тобто включення кори головного мозку в регуляцію функцій нижележащих структур мозку. Це дозволяє організувати більш досконалу, що враховує індивідуальний досвід, збережений у пам’яті, регуляцію функцій організму. Надалі, у міру автоматизації тієї чи іншої реакції, виконання її знову передається підкірковим структурам з формуванням автоматичного реагування.
Різні коркові зони дозрівають нерівномірно. Найбільш рано дозріває соматосенсорная і рухова кора, дещо пізніше – зорова і слухова. Особливо інтенсивним протягом першого півріччя життя є розвиток зорової кори, що тягне за собою розвиток інших зон мозку та їх інтеграцію. Дозрівання сенсорних і моторних зон в основному закінчується до 3 років. Значно пізніше дозріває асоціативна кора: до 7 років формуються її основні зв’язки, а остаточна диференціювання, формування нейронних ансамблів і зв`язків з іншими відділами мозку відбуваються до підліткового віку. Найбільш пізно (ближче до 9 років) дозрівають лобні області кори. Поступовість дозрівання структур кори великих півкуль визначає вікові особливості вищих нервових функцій і поведінкових реакцій дітей різних вікових груп.
Загальна площа кори головного мозку людини близько 2200 см2, число нейронів кори перевищує 10 млрд. У складі кори маються пірамідні, зірчасті, веретеноподібні нейрони.
Пірамідні нейрони мають різну величину, аксон пірамідного нейрона, як правило, проходить через білу речовину в інші зони кори або в інші мозкові структури.
Зірчасті клітини мають короткі добре гілкуються дендрити і короткий аксон, що забезпечує зв’язки нейронів в межах самої кори головного мозку.
Веретеноподібні нейрони забезпечують вертикальні або горизонтальні взаємозв’язку нейронів різних шарів кори.
Кора головного мозку має переважно шестіслойних будову (рис. 11.10).
Рис. 11.10. Будова кори головного мозку
Шар I – верхній молекулярний, представлений в основному ветвлениями висхідних дендритів пірамідних нейронів, серед яких розташовані рідкісні горизонтальні клітини і клітини-зерна, сюди ж приходять волокна неспецифічних ядер таламуса, що регулюють через дендрити цього шару рівень збудливості кори головного мозку.
Шар II – зовнішній зернистий, складається з зірчастих клітин, що визначають тривалість циркулювання збудження в корі головного мозку, тобто мають відношення до пам’яті.
Шар III – зовнішній пірамідний, формується з пірамідних клітин малої величини і разом з II шаром забезпечує Корко-коркові зв’язку різних звивин мозку.
Шар IV – внутрішній зернистий, містить переважно зірчасті клітини. Тут закінчуються специфічні таламокортікальние шляху, тобто шляху, що починаються від рецепторів аналізаторів.
Шар V – внутрішній пірамідний (гангліозний), шар великих пірамід, які є вихідними нейронами, аксони їх йдуть в стовбур мозку і спинний мозок. У руховій зоні в цьому шарі знаходяться гігантські пірамідні клітини, відкриті Бецом (клітини Беца).
Шар VI – шар поліморфних клітин, більшість нейронів цього шару утворюють кортико-таламические шляху.
Розподіл нейронів по шарах в різних областях кори дозволило виділити в мозку людини 53 цитоархитектонических поля (поля Бродмана), які вдосконалюються в міру розвитку кори головного мозку. У людини і вищих ссавців розрізняють, поряд з первинними, вторинні і третинні коркові поля, що забезпечують асоціацію функцій даного аналізатора з функціями інших аналізаторів.
Особливістю коркових полів є екранний принцип їх функціонування, що полягає в тому, що рецептор проектує свій сигнал не так на один нейрон кори, а на поле нейронів, яке утворюється їх зв’язками. У результаті сигнал фокусується не крапка в точку, а на безлічі різноманітних нейронів, що забезпечує його повний аналіз і можливість передачі в інші зацікавлені структури. Так, одне волокно, що приходить в зорову область кори, може активувати зону розміром 0,1 мм. Це означає, що один аксон розподіляє свою дію на більш ніж 5000 нейронів.
Функції окремих зон нової кори визначаються особливостями її структурної організації, зв’язками з іншими структурами мозку, участю в сприйнятті, зберіганні та відтворенні інформації при організації та реалізації поведінки, регуляції функцій сенсорних систем, внутрішніх органів.
Структурний відмінність ділянок кори головного мозку пов’язане з різницею їхніх функцій. У корі головного мозку виділяють сенсорні, моторні і асоціативні області (рис. 11.11).
Рис. 11.11. Цітоархітектоніческі поля Бродмана
Коркові кінці аналізаторів мають свою топографію – локальне розташування в певних ділянках кори головного мозку. Вони називаються сенсорними областями кори головного мозку. Коркові кінці аналізаторів різних сенсорних систем перекриваються. Крім цього, у кожній сенсорній системі кори маються полісенсорній нейрони, які реагують не тільки на “свій” адекватний стимул, але й на сигнали інших сенсорних систем. Ці механізми лежать в основі формування полімодальних зв’язків, що забезпечують сочетанную реакцію на різні подразники.
Шкірна рецептірующая система, таламокортікальние шляху проектуються на задню центральну звивину. Тут мається суворе соматотопическую поділ. На верхні відділи цієї звивини проектуються рецептивні поля шкіри нижніх кінцівок, на середні – тулуба, на нижні відділи – руки, голови.
На задню центральну звивину в основному проектується больова і температурна чутливість. У корі тім’яної частки (поля 5 і 7, див. Рис. 11.11), де також закінчуються провідні шляхи чутливості, здійснюється більш складний аналіз: локалізація роздратування, дискримінація, стереогіоз. При пошкодженнях кори особливо сильно порушуються функції дистальних відділів кінцівок, особливо рук.
Зорова система знаходиться в потиличній частці мозку: поля 17, 18, 19. Центральний зоровий шлях закінчується в поле 17; він інформує про наявність та інтенсивності зорового сигналу. У полях 18 і 19 аналізуються колір, форма, розміри, якість предметів. Поразка поля 19 кори головного мозку призводить до того, що хворий бачить, але не впізнає предмет (зорова агнозія, при цьому втрачається також колірна пам’ять).
Слухова система проектується в поперечних скроневих звивинах (звивини Гешля), в глибині задніх відділів латеральної (Сільвієвій) борозни (поля 41, 42, 52). Саме тут закінчуються аксони задніх горбів Четверохолміе і латеральних колінчастих тіл.
Нюхова система проектується в області переднього кінця гіппокампальних звивини (поле 34). Кора цій області має не шести-, а тришарове будову. При подразненні відзначаються нюхові галюцинації, пошкодження її веде до аносмія (втрати нюху).
Смакова система проектується в гіппокампальних звивині по сусідству з нюхової областю кори (поле 43).
У передній центральній звивині розташовані зони, подразнення яких викликає рух, вони представлені по соматотопическую типу, але зовсім інакше: у верхніх відділах звивини – нижні кінцівки, в нижніх – верхні. Це моторні області кори головного мозку.
Спереду від передньої центральної звивини лежать премоторні поля 6 і 8. Вони організовують не ізольовані, а комплексні, координовані, стереотипні рухи. Ці поля також забезпечують регуляцію тонусу гладкої мускулатури, пластичний тонус м’язів через підкіркові структури.
У реалізації моторних функцій беруть участь також друга лобова звивина, потилична, верхнетеменная області.
Рухова область кори як ніяка інша має велику кількість зв’язків з іншими аналізаторами, чим, мабуть, і зумовлена наявність в пий значного числа полісенсорних нейронів.
Всі сенсорні проекційні зони і моторні області кори займають менше 20% поверхні кори головного мозку. Решта – це асоціативні області. Кожна асоціативна область кори пов’язана потужними зв’язками з кількома проекційними областями. У асоціативних областях відбувається інтеграція разномодальних інформації, що дозволяє здійснювати усвідомлення надходить інформації та складні поведінкові акти. Асоціативні області мозку у людини найбільш виражені в лобової, тім’яної і скроневій.
Кожна проекційна область кори оточена асоціативними областями. Нейрони цих областей здатні до сприйняття разномодальних інформації, володіють великими здібностями до навчання. Полісенсорній нейронів асоціативної області кори забезпечує їх участь в об’єднанні, що надходить, забезпеченні взаємодії сенсорних і моторних областей кори.
Так, в тім’яній асоціативної області кори формуються суб’єктивні уявлення про навколишній простір, про нашому тілі. Це стає можливим завдяки зіставленню соматосенсорной, пропріоцептивної і зорової інформації. Лобові асоціативні поля мають зв’язку з лимбичним відділом мозку і беруть участь в організації програм дії при реалізації складних поведінкових актів з урахуванням їх емоційного забарвлення.
Першою і найбільш характерною рисою асоціативних областей кори є здатність їх нейронів сприймати разномодальную інформацію, причому сюди надходить не первинна, а вже оброблена інформація з виділенням біологічної значущості сигналу. Це дозволяє формувати програму цілеспрямованої поведінкового акту.
Друга особливість асоціативної області кори полягає в здатності до пластичних перебудовам в залежності від значимості надходить.
Третя особливість асоціативної області кори виявляється у тривалому зберіганні слідів сенсорних впливів. Руйнування асоціативної області призводить до виражених порушень навчання, пам’яті.
Розподіл функцій по областях мозку не є абсолютним. Встановлено, що практично всі області мозку мають полісенсорній нейрони, які певною мірою можуть брати на себе функцію пошкоджених модальноспсціфічсскіх нейронів. Це дозволяє компенсувати пошкодження структур мозку в ті періоди дитинства, коли пошкоджена функція ще не закріплена жорстко в структурі нервової тканини.
Важливою особливістю кори головного мозку є її здатність тривало зберігати сліди збудження. Це властивість надає корі виняткове значення в механізмах асоціативної переробки та зберігання інформації, накопичення знань.
Источник