Зон кори головного мозку

Окремі ділянки кори мають різне функціональне значення. Разом з підкірковими центрами, стовбуром мозку і спинним мозком великий мозок об’єднує окремі частини організму в єдине ціле, здійснює нервову регуляцію всіх органів (рис. 55, 56).
У кору великого мозку надходять доцентрові імпульси від рецепторів. Кожному рецепторному апарату відповідає в корі ділянка, яку І.П Павлов назвав кірковим ядром аналізатора. Ділянка кори, де розташовані кіркові ядра аналізаторів, названі сенсорними зонами кори великого мозку.
Ядерна зона рухового аналізатора (сомато-сенсорна зона), куди надходять збудження від рецепторів суглобів, скелетних м’язів і сухожилок, розташована в передню- і задньоцентральних ділянках кори. У межах передньої центральної закрутки найвище розміщені центри для м’язів нижньої кінцівки, нижче – для м’язів тулуба, потім верхньої кінцівки і, нарешті, центри м’язів голови. Ураження цієї зони призводить до паралічу протилежної половини тіла.
Рис. 55. Зовнішня будова головного мозку (за дорлінг кіндерслі, 2003)
Передцентральна звивина (gyrus precentralis) і прицентральна часіка (lobulus paracentralis) лобової частки становлять руховий центр кори і є аналізатором кінестезичних імпульсів, які надходять від посмугованих м’язів, суглобів, сухожилків. Тут замикаються рухові умовні рефлекси. У верхній ділянці перед центральної звивини розташовані клітинні групи, що належать до м’язів нижніх кінцівок, нижче – верхніх кінцівок, ще нижче – неврони, пов’язані з іннервацією м’язів голови. Оскільки нервові шляхи перехрещуються, праві рухові центри кори пов’язані з мускулатурою лівої сторони тіла і навпаки.
У задній частині середньої лобової звивини міститься центр узгодженого руху голови й очей (окоруховий, блоковий, відвідний і додатковий нерви).
Рис. 56. Кіркові поля (за Дорлінг Кіндерслі, 2003)
У задньому відділі нижньої лобової звивини розміщена зона Брока – руховий центр мови, який разом із центром Верніке забезпечують здатність людини читати, писати, чути, вимовляти і розуміти мову.
Ушкодження різних полів кори лобової частки може призвести: до підвищення агресивності й послаблення реакції страху; зростання пасивно-захисних умовних рефлексів; порушення харчових і захисних умовних рефлексів.
У таких людей спостерігається втрата ініціативи, апатія, порушення абстрактного мислення, нездатність до творчого мислення, розгальмування нижчих емоцій і потягів, розлади мовлення і понятійного мислення.
У задній частині лобової звивини розташований центр письма, ураження якого призводить до порушення навичок письма під контролем зору.
У лівій (у лівшів у правій) нижній тім’яній часточці розташований центр, який координує цілеспрямовані рухи. Він функціонує за типом тимчасових зв’язків, які виникають протягом індивідуального життя, тобто умовних рефлексів. У разі ушкодження цього центру елементи довільних рухів зберігаються, але порушуються цілеспрямовані дії (апраксія).
У верхній тім’яній частці (задньоцентральна звивина) розміщений кірковий центр аналізаторів чутливості (больової, температурної, дотикової), або сомато-сенсорна кора. Ураження кори у цій частині призводить до часткової або повної анестезії (втрата чутливості).
Ураження кори в ділянці верхньої тім’яної частки призводить до зниження больової чутливості і порушення стереогноза – впізнавання предметів на дотик без допомоги зору.
У нижній тім’яній частині розташований центр праксії, який регулює здатність здійснювати координаційні рухи, які складають основу робочих процесів і потребують спеціального навчання.
У кутовій звивині тім’яної частки розташований зоровий центр мови. Його ураження призводить до неможливості розуміння письма (алексія).
Тім’яна ділянка – це апарат вищої інтегративної діяльності мозку людини, вона безпосередньо стосується процесів біологічної і соціальної адаптації, є фізіологічною основою вищих психічних функцій.
Локалізація статичного аналізатора (центр збереження рівноваги і положення тіла в просторі) – кора верхньої та середньої скроневих звивин. Ушкодження цього центру призводить до атаксії (розладу координації рухів).
Зона шкірного аналізатора, зв’язаного з температурою, больовою і тактильною чутливістю займає задньоцентральну ділянку. Центри чутливості нижчих частин тіла розміщені у верхніх частинах тіла – у нижніх її ділянках.
Найбільшу площу займає кіркове представництво рецепторів кисті рук, голосового аналізатора і обличчя, найменшу – тулуба, стегна і гомілки.
Ядерна зона зорового аналізатора розташована на внутрішній поверхні потиличної ділянки, в зоні шпорної борозни. Ураження цього центру призводить до сліпоти. При порушеннях у сусідніх із шпорною борозною частин кори в ділянці потиличного полюса на медіальній і латеральній поверхнях частки може спостерігатися втрата зорової пам’яті, здатності орієнтації у незнайомій обстановці, порушення функції, пов’язаної із бінокулярним зором (здатності за допомогою зору оцінювати форму предметів, відстань до них тощо).
У корі верхньої скроневої звивини розташована частина слухового аналізатора, а поблизу від бокової борозни – ядерна зона смакового аналізатора. Двостороннє ураження до повної кіркової глухоти.
Нюхова зона розміщена на внутрішній поверхні скроневих ділянок кори. В ділянці середньої і нижньої скроневих звивин розташоване кіркове представництво вестибулярного аналізатора. Ураження цієї ділянки призводить до порушення рівноваги під час стояння і зниження чутливості.
Із сенсорними зонами взаємодіє моторна зона кори великого мозку. Ядерні зони аналізаторів – це ділянки кори, в яких закінчується основна маса провідних шляхів аналізаторів. За межами ядерних зон розташовані розсіяні елементи, куди надходять імпульси від тих же рецепторів, що і в ядро аналізатора.
Центр мови міститься у лівій півкулі. Розрізняють 2 центри мови: руховий (зона Брока), який міститься у нижній частині лобової ділянки і слуховий (зона Верніке), який знаходиться у скроневій ділянці, під заднім кінцем сільвієвої борозни. Центри мови є лише у людини. Мовлення, мислення, почуття і вправні рухи контролюються нейронами, які розміщені в лобовій ділянці головного мозку. Розпізнавання тонів і звуків відбувається в скроневій ділянці. Ця ділянка також бере участь у запам’ятовуванні інформації. Різноманітні сенсорні відчуття, такі як біль, температура усвідомлюються та інтерпретуються в тім’яній ділянці. Потилична ділянка фіксує та інтерпретує зорові образи.
Источник
Кора большого мозга представлена равномерным слоем серого вещества толщиною 1,3-4,5 мм, состоящим более чем из 14 млрд. нервных клеток. Благодаря складчатости коры ее поверхность достигает больших размеров — около 2200 см2.
Толща коры состоит из шести слоев клеток, которые различают при специальной окраске и исследовании под микроскопом. Клетки слоев различны по форме и размерам. От них в глубь мозга отходят отростки.

Строение коры головного мозга
Было установлено, что разные участки — поля коры полушарий различаются по строению и функциям. Таких полей (называемых еще зонами, или центрами) выделяют от 50 до 200. Строгих границ между зонами коры большого мозга не существует. Они составляют аппарат, обеспечивающий прием, переработку приходящих сигналов и ответную реакцию на поступившие сигналы.
Зоны коры головного мозга
В задней центральной извилине, позади от центральной борозды, располагается зона кожной и суставно-мышечной чувствительности. Здесь воспринимаются и анализируются сигналы, возникающие при касании к нашему телу, при воздействии на него холода или тепла, болевых воздействиях.
 Зоны коры головного мозга
Зоны коры головного мозга
В противоположность этой зоне — в передней центральной извилине, спереди от центральной борозды, расположена двигательная зона. В ней выявлены участки, которые обеспечивают движения нижних конечностей, мышц туловища, рук, головы. При раздражении этой зоны электротоком возникают сокращения соответствующих групп мышц. Ранения или другие повреждения коры двигательной зоны влекут за собой паралич мышц тела.
В височной доле находится слуховая зона. Сюда поступают и здесь анализируются импульсы, возникающие в рецепторах улитки внутреннего уха. Раздражения участков слуховой зоны вызывают ощущения звуков, а при поражении их болезнью утрачивается слух.
Зрительная зона расположена в коре затылочных долей полушарий. При ее раздражении электрическим током во время операций на мозге человек испытывает ощущения вспышек света и темноты. При поражении ее какой-либо болезнью ухудшается и теряется зрение.
Вблизи боковой борозды расположена вкусовая зона, где анализируются и формируются ощущения вкуса на основании сигналов, возникающих в рецепторах языка. Обонятельная зона расположена в так называемом обонятельном мозге, у основания полушарий. При раздражении этих зон во время хирургических операций или при воспалении люди ощущают запах или вкус каких-либо веществ.
Чисто речевой зоны не существует. Она представлена в коре височной доли, нижней лобной извилине слева, участках теменной доли. Их поражения болезнями сопровождаются расстройствами речи.
Первая и вторая сигнальные системы
Неоценима роль коры большого мозга в совершенствовании первой сигнальной системы и развитии второй. Эти понятия разработаны И.П.Павловым. Под сигнальной системой в целом понимают всю совокупность процессов нервной системы, осуществляющих восприятие, переработку информации и ответную реакцию организма. Она связывает организм с внешним миром.
Первая сигнальная система
Первая сигнальная система обусловливает восприятие посредством органов чувств чувственно-конкретных образов. Она является основой для образования условных рефлексов. Эта система существует как у животных, так и у человека.
В высшей нервной деятельности человека развилась надстройка в виде второй сигнальной системы. Она свойственна только человеку и проявляется словесным общением, речью, понятиями. С появлением этой сигнальной системы стали возможными отвлеченное мышление, обобщение бесчисленных сигналов первой сигнальной системы. По И.П.Павлову, слова превратились в «сигналы сигналов».
Вторая сигнальная система
Возникновение второй сигнальной системы стало возможным благодаря сложным трудовым взаимоотношениям между людьми, так как эта система является средством общения, коллективного труда. Словесное общение не развивается вне общества. Вторая сигнальная система породила отвлеченное (абстрактное) мышление, письмо, чтение, счет.
Слова воспринимаются и животными, но совершенно отлично от людей. Они воспринимают их как звуки, а не их смысловое значение, как люди. Следовательно, у животных нет второй сигнальной системы. Обе сигнальные системы человека взаимосвязаны. Они организуют поведение человека в широком смысле слова. Причем вторая изменила первую сигнальную систему, так как реакции первой стали в значительной мере зависеть от социальной среды. Человек стал в состоянии управлять своими безусловными рефлексами, инстинктами, т.е. первой сигнальной системой.
Функции коры мозга
Знакомство с наиболее важными физиологическими функциями коры большого мозга свидетельствует о необычайном ее значении в жизнедеятельности. Кора вместе с ближайшими к ней подкорковыми образованиями является отделом центральной нервной системы животных и человека.
Функции коры головного мозга — осуществление сложных рефлекторных реакций, составляющих основу высшей нервной деятельности (поведения) человека. Не случайно у него она получила наибольшее развитие. Исключительным свойством коры являются сознание (мышление, память), вторая сигнальная система (речь), высокая организация труда и жизни в целом.
Источник
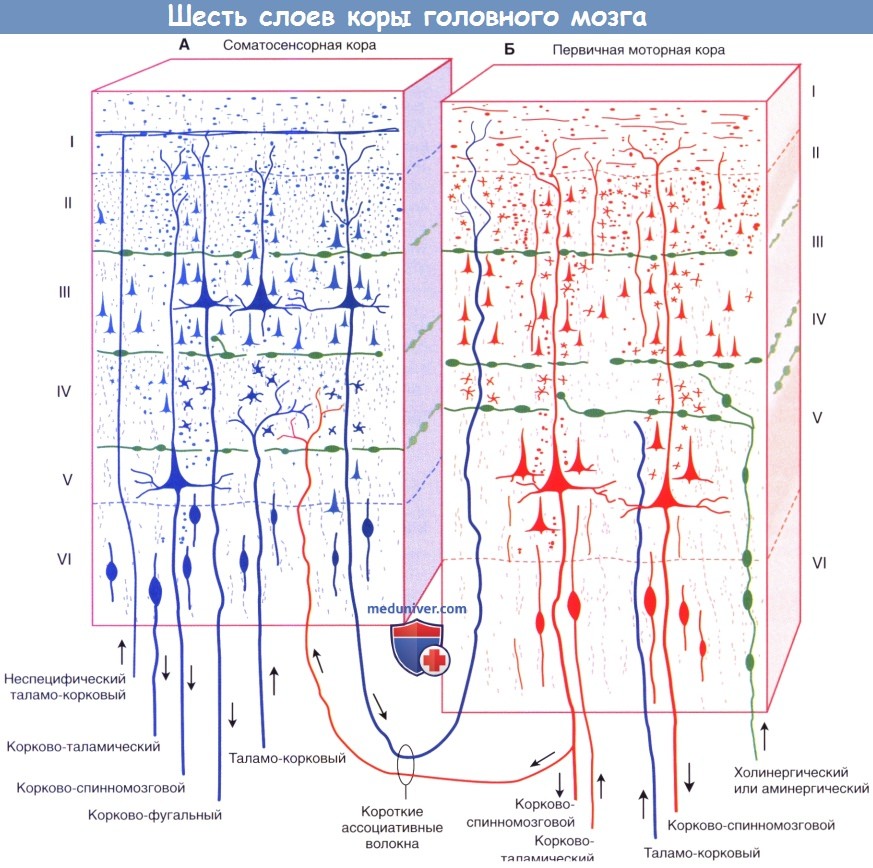
Строение коры головного мозга: слои, клеткиТолщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть. Микроскопически кора имеет полосчатый, или пластинчатый, вид, отражающий распределение ее клеток и нервных волокон, а также веерообразную организацию клеточных элементов. Общая цитоархитектоника (схема строения, основанная на типе клеток; схему строения в зависимости от распределения миелинизированных волокон обозначают термином миелоархитектоника) во многом варьирует от зоны к зоне, что позволяет составить «карту» коры с десятками гистологически различных зон. Больших успехов удалось достичь в соотнесении этих зон со специфическими функциями. Несмотря на то, что это имеет теоретическую ценность, подобное разделение довольно упрощено, так как часто принимают во внимание только ключевые функции более обширных функциональных систем, взаимодействующих с другими отделами мозга. а) Пластинчатое строение. Пластинчатое распределение нейронов характерно для всех отделов коры. Филогенетически «старые отделы», включая древнюю кору (обонятельную кору) и старую кору (гиппокампальная формация и зубчатое ядро; участвуют в процессах памяти) образованы тремя клеточными пластинками. Однако в новой коре (неокортексе или изокортексе, название которой указывает на единый нейрогенез коры, закончившийся образованием шестислойного строения), составляющей оставшиеся 90% (подавляющую часть коры больших полушарий), происходит переход к шестислойному строению.
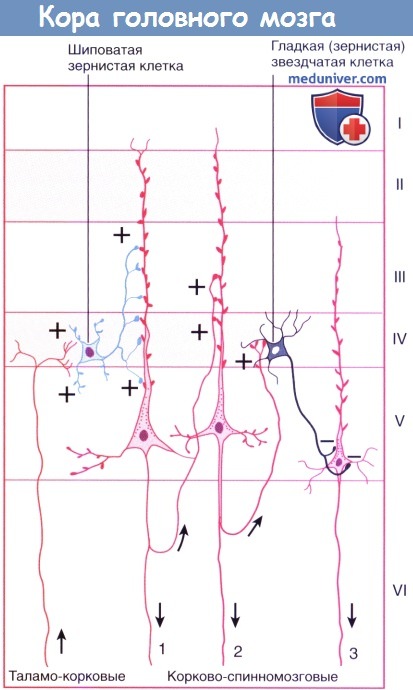
Клеточные пластинки новой коры (неокортекса): I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса. II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками. III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток. IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора. V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу. VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу. б) Колончатое строение. Помимо хорошо различимой пластинчатой организации коры больших полушарий, можно наблюдать также лучистую, или «колончатую», организацию клеточных структур. Колончатое строение новой коры стало основой исследования функционирования групп нейронов в соматосенсорной коре животных. Подобные лучевидно расположенные группы клеток, вероятно, представляют собой отдельные зоны с одинаковыми физиологическими свойствами и образуют структурную основу для создания более сложных функций. Группы колонн могут образовывать модули, участвующие в обработке различных сенсорных модальностей и функций. Установлено, что колонны в различных отделах коры имеют разное строение, обусловленное вариабельностью многочисленных свойств, включая строение и число клеток, онтогенез, синаптические связи и молекулярные маркеры. Все это влияет на различные функциональные характеристики и реакции в ответ на возбуждение. В качестве организующего принципа подобная концепция колончатого строения представляет определенную пользу, однако удобнее рассматривать кору как совокупность горизонтального (пластинчатого) и вертикального (лучистого) типов строения. Несмотря на то, что колонны не представляют собой отдельные подобные образования с четкими границами, такая концепция более точно отражает строение коры, а также наблюдаемые экспериментально функциональность, «экономию пространства» и пластичность коры. Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи. Лежащая в основе организации коры «схема» приводит к тому, что каждая из колонн становится модально (функционально) специфичной после «обработки» информации отдельными элементами. Однако конечная реакция проекционных нейронов колонн может значительно варьировать в зависимости от степени их возбуждения и информации, подходящей к каждому нейрону. Например, одна колонна может реагировать на движение в определенном суставе, но не на раздражение кожи над ним; однако при изменении условий их реакция также может изменяться.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический. Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.) • Пирамидные клетки имеют пирамидоподобную форму с вершиной, направленной к поверхности. Размеры клеток в высоту составляют 20-30 нм в слоях II и III и более чем в два раза больше в слое V. Самые крупные — гигантские пирамидные клетки Беца — расположены в моторной коре. Единственный апикальный дендрит каждой пирамидной клетки достигает слоя I, заканчиваясь на пучке дендритов. Несколько базальных дендритных ветвей отходят от базальных «углов» клетки и веерообразно расходятся к соответствующим слоям. Апикальные и базальные дендритные ветви свободно разветвляются и усыпаны дендритными шипиками. Большинство пирамидных клеток расположено в слоях II-III и V-VI. Отходящие от основания клеток аксоны отдают несколько возвратных ветвей до вхождения в подлежащее белое вещество, их функция — возбуждение соседних пирамидных клеток. • Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением. Нейромедиатором различных по структуре и классификации непирамидных тормозных вставочных нейронов служит ГАМК. [Сложная классификация нейронов новой коры постоянно меняется. Гладкие звездчатые (зернистые) клетки можно обнаружить во всех слоях коры; их дендриты веерообразно расходятся во всех направлениях, а их аксоны образуют локальные древовидные разветвления, поэтому их иногда называют нейронами локальных сплетений. Несмотря на их уникальные морфологические характеристики, нейроглиеформные, клетки-канделябры и корзинчатые клетки считают специализированными типами зернистых клеток. Наш совет: если Вы встречаете термины «зернистая» или «гладкая звездчатая» клетка, для облегчения чтения и понимания их следует рассматривать как вставочные нейроны.] Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR). • Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях. Корзинчатые клетки лежат преимущественно в слоях II и V, а свое название получили благодаря тому, что их аксоны формируют околоклеточные «корзинки» вокруг тел пирамидных клеток, дистальных отделов их дендритов и аксонов других корзинчатых клеток. • Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств. • 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах. 1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария. 2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария. 3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках. 4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы: д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга. • Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III. • Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III. • Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса). – Также рекомендуем “Корковые зоны головного мозга: цитоархитектонические поля Бродмана, методы исследования” Редактор: Искандер Милевски. Дата публикации: 22.11.2018 |
Источник