Рухова зона кори головного мозку

Двигательная область коры головного мозга: расположение, строение и функции
Часть мозга, которая выполняет работу произвольных движений является двигательной (моторной) корой. Необходимые движения осуществляются таким образом, чтобы они лучше всего подходят для текущей позиции индивида.
Левая двигательная кора участвует в регулировании движения правой стороны тела, а правая двигательная область коры головного мозга координирует движения левой стороны тела.
Задумывались ли вы, какая часть мозга играет решающую роль в обеспечении произвольных движений? Это двигательная кора. Она расположена в задней части лобной доли ― 1 из 4 основных долей, передней части мозга. Двигательная кора лежит перед центральной бороздой, как структура, которая позволяет отличить лобную долю и теменной доли.
Функция
Как следует из названия, двигательная кора отвечает за двигательные функции тела, она принимает участие в движении мышц для выполнения определенных задач. Наша способность производить движения, такие как собирание мелких предметов, привязаны к нормальной работе двигательной коры.
Будь то небольшие движения, такие как перемещение пальцев или большие движения, такие, как ходьба и бег, двигательная кора играет решающую роль прямо от начала выполнения движения. Проще говоря, сокращение мышц для нормального движения – это то, что находится под контролем двигательной коры.
В момент начала движения, верхние двигательные нейроны, расположенные в двигательной коре, передают электрические сигналы, которые идут через другие структуры мозга, такие как базальные ганглии и мозговой ствол, и поступают на нижние моторные нейроны, расположенные в спинном мозге. Сигнал затем проходит вниз от нижнего моторного нейрона к другому и, наконец, прибывает на нужные группы мышц. Получив сигнал, мышцы сокращаются, вызывая произвольные движения.
Премоторная кора
Для того, чтобы выполнить определенное движение, определенные мышцы должны сужаться. Премоторная кора головного мозга анализирует и точно определяет группы мышц, которые должны быть использованы для выполнения определенных движений. Она учитывает текущее положение и позу тела при выборе набора мышц для выполнения движения. Например, пиная футбольный мяч, премоторная кора решает какие мышцы ног задействовать, чтобы инициировать действие.
Первичная моторная кора
Эта часть мозга непосредственно взаимодействует с двигательными нейронами спинного мозга, чтобы стимулировать предназначенные мышцы. Короче говоря, первичная моторная кора участвует в передаче сигналов, которые вносят непосредственный вклад в создание движения.
Дополнительная моторная область
Одна из основных функций деятельности ДМО является надзор за ручной координацией. В ручной координации, множественные движения должны быть выполнены одновременно для достижения желаемого действия или задачи.
Например, действия, которые предполагают использование обеих рук потребуют координации между левой и правой двигательной корой. Это делается ДМО, которая позволяет производить совмещение движений одновременно для выполнения сложных действий. Поэтому наша способность двигать одновременно обе руки для выполнения определенной задачи приписывается работу ДМО.
ДМО также помогает определять последовательность движений, так как для выработки оптимального пути движения, необходимо осуществлять комплекс действий.
Сразу отмечу еще один момент заключается в том, что двигательная кора не решает, выполнять ли движение или нет. Например, увидев кусочек шоколада, лежащего на полу, решение его забрать принимается префронтальной корой, которая лежит непосредственно перед премоторной. Префронтальная кора, соответственно, дает сигналы двигательной области коры головного мозга, чтобы осуществлять движение. Кроме того, двигательная кора не единственная область, которая участвует в выполнении точных движений. Другие части мозга, такие как базальные ганглии и мозжечок, в равной степени способствуют производству движений.
Источник
Моторные зоны коры головного мозга: моторная кора, премоторная кора, корковые глазодвигательные поля
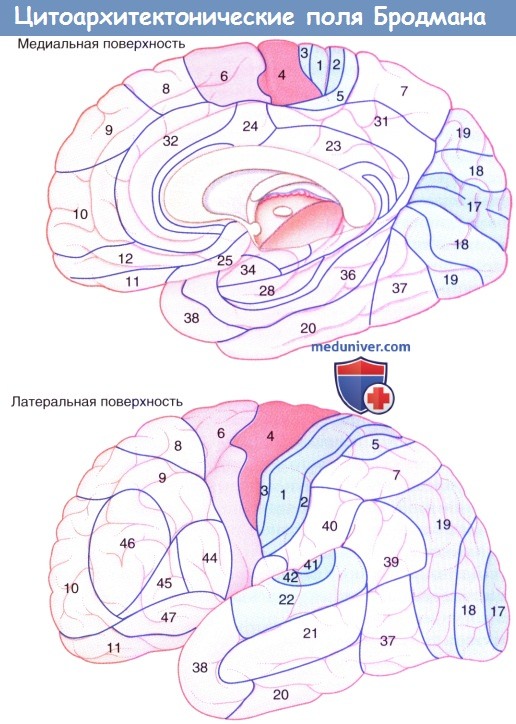
а) Первичная моторная кора. Первичная моторная кора (поле 4) — полоска агранулярной коры в прецентральной извилине. Она дает начало 60-80% (количество варьирует) волокон корково-спинномозгового пути (КСП). Остальная часть волокон отходит от премоторной, поясной и добавочной моторной зон, а также теменной коры, что показано в отдельной статье на сайте. Наибольшую плотность окончаний КСП в спинном мозге наблюдают в зонах, иннервирующих дистальные мышцы конечностей. Части тела противоположной стороны соматотопически представлены в коре инвертированными, за исключением лица, а относительно крупные области занимают кисть (играет важную роль в контроле движений пальцев), околоротовая область и язык. Зону кисти обычно можно определить по направленному кзади возвышению в 6-7 см от верхнего края полушария. Ипсилатералъные части тела также имеют соматотопическую организацию; ипсилатеральные моторные нейроны составляют лишь 10% волокон пирамидного пути, которые не переходят на противоположную сторону и не иннервируют дистальные мышцы конечностей. При целенаправленной стимуляции моторной коры человека установлено, что клеточные колонны регулируют направление движения. Первичная моторная кора «синтезирует» двигательные команды, но не является местом их возникновения. От нее волокна в составе КСП направляются в спинной мозг и широко разветвляются по мере достижения места своего окончания. Для того чтобы взять, например, ручку в руку, требуется: Удержание всей руки при любом типе манипуляций — функция премоторной коры, отражающая важность бессознательной регуляции положения тела при совершении произвольных движений. Более крупные моторные зоны коры формируются при «объединении» близлежащих нейронных колонн на основе их функций и создании сложных последовательностей движений. 1. Пластичность моторной коры. Некрупные поражения моторной коры у обезьян и низших млекопитающих приводят первоначально к параличу соответствующей части тела, а затем в течение нескольких дней (иногда часов) — к быстро прогрессирующему восстановлению функции. Восстановление может быть обусловлено изменением свойств клеточных колонн, расположенных рядом с местом поражения, которые берут на себя утраченную двигательную функцию. Увеличение двигательной зоны участка коры возможно путем локального введения антагонистов ГАМК в кору. Расширение моторной зоны на уровне спинного мозга заранее обеспечено обширными зонами перекрытия от поля 4 до колонок двигательных нейронов серого вещества переднего рога, однако степень пластичности здесь ниже, чем в коре. Взаимосвязи между КСП (и другими нисходящими путями) и двигательными нейронами в спинном мозге возникают посредством вставочных нейронов. Во вставочных нейронах происходит объединение сенсорной и корковой информации, что сопровождается специфической и организованной активацией пулов двигательных нейронов и сокращением иннервируемых ими мышц. 2. Источники афферентных путей к первичной моторной коре:
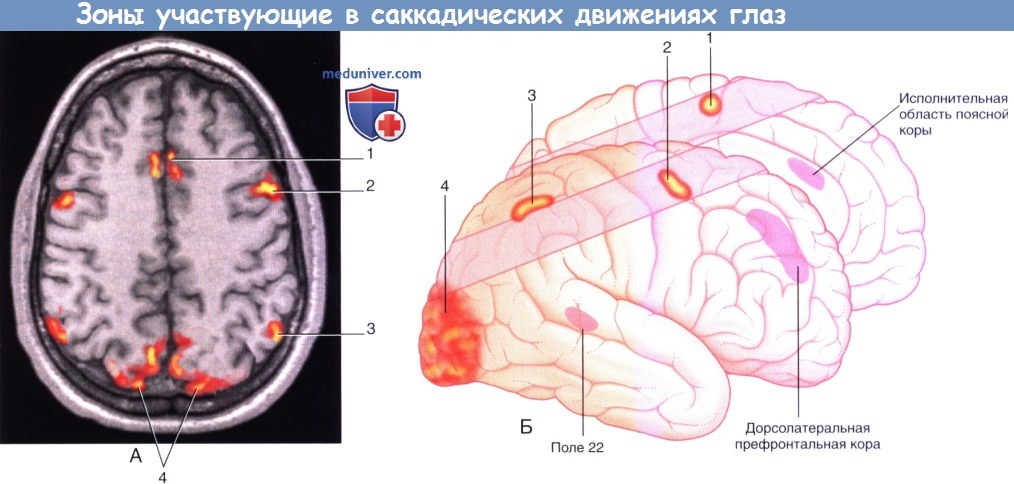
б) Премоторная кора. Премоторная кора (поле 6 на латеральной поверхности полушария) примерно в 6 раз крупнее первичной моторной коры. Она получает когнитивные импульсы от лобной доли при намерении произвести движение, а также от теменной доли (поле 7), передающей огромное количество тактильной и пространственно-зрительной информации. Максимальная ее активация происходит, когда последовательность движений происходит в ответ на зрительные или соматосенсорные раздражители, такие как приближение к объекту, полностью находящему в поле зрения, или идентификация объекта, находящегося вне поля зрения путем ощупывания. Активация премоторной коры, как правило, двусторонняя. Одним из объяснений служит межполушарная передача двигательных программ через мозолистое тело. Следует также принимать во внимание обширные связи премоторной коры с ядрами ствола мозга, дающими начало ретикуло-спинномозговым путям (и небольшому пути к КСП). Поражения, затрагивающие премоторную кору у человека, встречают редко, они характеризуются нестабильностью положения в противоположном плече и бедре. Таким образом, важной функцией премоторной коры, вероятно, служит двустороннее поддержание позы, например фиксация плечей при работе двумя руками и стабилизация бедер при ходьбе. Премоторная кора может вносить вклад в восстановление функции при истинной моторной гемиплегии, возникающей вследствие сосудистого поражения, затрагивающего КСП в лучистом венце. При данном поражении во время ПЭТ-исследования наблюдают повышение активности премоторной коры; кпереди от КСП спускается корково-ретикуло-спинномозговой проводящий путь. в) Дополнительная моторная зона (ДМЗ). В отличие от премоторной коры, реагирующей на внешние раздражители, ДМЗ (поле 6 на медиальной поверхности полушария) отвечает на внутренние раздражители и принимает особое участие в планировании движений. Это подтверждает тот факт, что ДМЗ возбуждается лобной долей (ДЛПФК) в тот момент, когда мы собираемся совершить движение, даже если движение в итоге не возникает. Важнейшей функцией ДМЗ, вероятно, является предварительное программирование последовательностей движений, уже заложенных в двигательной памяти. ДМЗ функционирует совместно с моторной петлей, проходящей через базальные ганглии, а также взаимодействует с полем 4 и отдает волокна непосредственно в КСП. Одностороннее поражение ДМЗ может сопровождаться акинезией (невозможностью начать движение) руки и ноги с разных сторон. Двустороннее поражение приводит к тотальной акинезии, включая речевую акинезию. 1. Корковые глазодвигательные поля. На рисунке ниже показаны корковые глазодвигательные поля, участвующие в сканирующих движениях (саккадах). Их взаимодействия и функции представлены в таблице ниже. 2. Дорсолатеральная префронтальная кора (ДЛПФК). ДЛПФК — высший когнитивный центр, участвует в оценке зрительной картины, принятии решения о сознательном совершении саккад и добровольном подавлении рефлекторных саккад (произвольные саккады возникают по воле человека). Рефлекторные саккады — автоматические реакции на объекты, появляющиеся в периферическом поле зрения. Строго говоря, рефлекторные саккады следует называть ответными; они не являются истинными рефлексами, так как способны подавляться волей. 3. Поясная кора. Вместе с ДЛПФК участвует в принятии решений и оценке эмоциональной важности, или ценности, видимых объектов. 4. Дополнительное глазодвигательное поле. Занимает передний отдел ДМЗ и участвует в планировании движений, особенно, если требуются множественные саккадические движения.
5. Фронтальное глазодвигательное поле. Фронтальное глазодвигательное поле (ФГДП) инициирует произвольные саккады, благодаря которым происходит переключение внимания на раздражитель или подавляется стремление к прямому взору на новый раздражитель в ответ на одно или более из трех перечисленных афферентных влияний. ФГДП «поддерживает» карту зрительного пространства относительно глазодвигательных координат и вместе с верхним холмиком играет ключевую роль в создании зрительно-направленных и произвольных саккад; поражения обеих структур приводят к необратимой утрате саккадических движений. Клинические и экспериментальные наблюдения у обезьян показали следующее. 6. Теменное глазодвигательное поле. Теменное глазодвигательное поле (ТГДП) отвечает за создание рефлекторных саккад и побуждает ФГДП к созданию произвольных саккад. ТГДП также участвует в пространственном восприятии, создавая схему зрительной картины. Поражения префронтальной коры и лобной доли описаны в отдельной статье на сайте.
в) Синдром ригидного человека (СРЧ). Редкое, но легко распознаваемое заболевание, известное как синдром ригидного человека (СРЧ), или синдром мышечной скованности, — аутоиммунное заболевание центральной нервной системы, связанное с наличием циркулирующих антител к глутаматдекарбоксилазе (GAD65),— ключевому ферменту, преобразующему глутамат в ГАМК. Синдром ригидного человека (СРЧ) проявляется скованностью мышц с эпизодическими мышечными спазмами (вызванными одновременным сокращением мышц синергистов и антагонистов, преимущественно в проксимальных отделах конечностей и осевой мускулатуре) и боязнью выполнения определенных задач. В норме контроль верхних моторных нейронов происходит за счет тонической активности прилежащих тормозных ГАМК-ергических вставочных нейронов. Одни зоны коры поражаются сильнее других, а клинические проявления связаны с нарушением функции этих ГАМК-ергических нейронов, что приводит к гипервозбудимости коры. В настоящее время уточняют роль циркулирующих антител в патогенезе синдрома ригидного человека (СРЧ). г) Резюме. Кора больших полушарий имеет одновременно пластинчатую и колончатую организацию. К двум основным типам клеток относят пирамидные и непирамидные клетки (вставочные нейроны). Пирамидные клетки расположены в слоях (пластинках) II, III, а также V (в виде веретеновидных клеток) и VI. Слой IV богат шиповатыми звездчатыми клетками (модифицированными пирамидными нейронами). Мелкие пирамидные клетки связывают извилины одного полушария, средние пирамидные клетки связывают между собой соответствующие зоны двух полушарий, а от крупных пирамидных клеток отходят волокна к таламусу, стволу мозга и спинному мозгу. Все корковые волокна считают возбуждающими; шиповатые звездчатые клетки также считают возбуждающими пирамидными клетками. Корковые вставочные нейроны—тормозные. Колончатая организация представлена колоннами клеток, которые рассматривают в качестве первичной функциональной единицы обработки информации в коре; они состоят из определенных нейронных «микроцепей». Части тела в соматосенсорной коре представлены инвертированными. Важная входящая информация поступает от вентрального заднего ядра таламуса, а направляется к первичной моторной коре и нижней теменной коре. В первичную зрительную кору входит коленчато-шпорный путь. Клеточные реакции различной сложности зависят от передачи информации с более простых типов клеток на более сложные. Свойство ассоциативных зрительных зон—распознавание черт, например цвета, формы и движения. Обработка цвета и формы продолжается в коре нижележащей височной доли, а движений—в задней теменной доле. Первичная слуховая кора занимает верхнюю поверхность верхней височной извилины, а ассоциативная слуховая кора расположена с латеральной стороны от нее. Первичная моторная кора занимает прецентральную извилину. Она дает начало большей части волокон пирамидного тракта, а части тела в ней представлены инвертированными. Основная информация подходит к ней от соматосенсорной коры, мозжечка (через вентральное заднее ядро таламуса), премоторной и дополнительной моторной зон. Премоторная кора отвечает преимущественно на внешние раздражители, а дополнительная моторная зона — на внутренние стимулы. Дорсолатеральная префронтальная кора контролирует четыре различных корковых зоны, в различной степени участвующих в создании противоположных саккад. – Также рекомендуем “Электроэнцефалография: нейрофизиологические основы, методика выполнения ЭЭГ” Редактор: Искандер Милевски. Дата публикации: 22.11.2018 |

Источник