Цикл кори и цикл кребса
Привет! Скоро лето, а значит, все второкурсники медвузов будут сдавать биохимию. Сложный предмет, на самом деле. Чтобы немного помочь тем, кто повторяет материал к экзаменам, я решил сделать статью, в которой расскажу вам о «золотом кольце» биохимии — цикле Кребса. Его также называют цикл трикарбоновых кислот и цикл лимонной кислоты, это всё синонимы.
Сами реакции я распишу в следующей статье. Сейчас же я расскажу о том, для чего нужен цикл Кребса, где он проходит и в чём его особенности. Надеюсь, получится понятно и доступно.
Для начала давайте разберём что такое обмен веществ. Это основа, без которой понимание Цикла Кребса невозможно.
Метаболизм
Одно из важнейших свойств живого (вспоминаем биологию) — это обмен веществ с окружающей средой. Действительно, только живое существо может что-то поглощать из окружающей среды, и что-то потом в неё выделять.
В биохимии обмен веществ принято называть «метаболизм». Обмен веществ, обмен энергией с окружающей средой — это метаболизм.
Когда мы, допустим, съели бутерброд с курицей, мы получили белки (курица) и углеводы (хлеб). В процессе пищеварения белки распадутся до аминокислот, а углеводы — до моносахаров. То, что я описал сейчас, называется катаболизм, то есть распад сложных веществ на более простые. Первая часть метаболизма — это катаболизм.
Ещё один пример. Ткани в нашем организме постоянно обновляются. Когда отмирает старая ткань, её обломки растаскивают макрофаги, и они заменяется новой тканью. Новая ткань создаётся в процессе синтеза белка из аминокислот. Синтез белка происходит в рибосомах. Создание нового белка (сложного вещества) из аминокислот (простого вещества) — это анаболизм.
Итак, анаболизм — это противоположность катаболизму. Катаболизм — это разрушение веществ, анаболизм — это создание веществ. Кстати, чтобы их не путать, запомните ассоциацию: «Анаболики. Кровью и потом». Это голливудский фильм (довольно скучный, на мой взгляд) о спортсменах, применяющих анаболики для роста мышц. Анаболики — рост, синтез. Катаболизм — обратный процесс.
Точка пересечения распада и синтеза.
Цикл Кребса как ступень катаболизма.
Как связаны метаболизм и цикл Кребса? Дело в том, что именно цикл Кребса является одной из важнейших точек, в которой сходятся пути анаболизма и катаболизма. Именно в этом и заключается его значение.
Давайте разберём это на схемках. Катаболизм можно условно представить как расщепление белков, жиров и углеводов в нашей пищеварительной системе. Итак, мы скушали пищу из белков, жиров, и углеводов, что дальше?
А дальше все эти вещества распадутся на простые составляющие:
- Жиры — на глицерин и жирные кислоты (могут быть и другие компоненты, я решил взять самый простой пример);
- Белки — на аминокислоты;
- Полисахаридные молекулы углеводов — на одинокие моносахариды.
Далее, в цитоплазме клетки, последует превращение этих простых веществ в пировиноградную кислоту (она же — пируват). Из цитоплазмы пировиноградная кислота попадёт в митохондрию, где превратится в ацетил коэнзим А. Пожалуйста, запомните эти два вещества — пируват и ацетил КоА, они очень важны.
Давайте теперь посмотрим, как происходит этап, который мы сейчас расписали:
Важная деталь: аминокислоты могут превращаться в ацетил КоА сразу, минуя стадию пировиноградной кислоты. Жирные кислоты сразу превращаются в ацетил КоА. Учтём это и подредактируем нашу схемку, чтобы получилось правильно:
Превращения простых веществ в пируват происходят в цитоплазме клеток. После этого пируват поступает в митохондрии, где успешно превращается в ацетил КоА.
Для чего пируват превращается в ацетил КоА? Именно для того, чтобы запустить наш цикл Кребса. Таким образом, мы можем сделать ещё одну надпись в схеме, и получится правильная последовательность:
В результате реакций цикла Кребса образуются важные для жизнедеятельности вещества, главные из которых:
- НАДH (НикотинАмидАденинДиНуклеотид+ катион водорода) и ФАДH2 (ФлавинАденинДиНуклеотид+молекула водорода). Я специально выделил заглавными буквами составные части терминов, чтобы легче было читать, в норме их пишут одним словом. НАДH и ФАДH2 выделяются в ходе цикла Кребса, чтобы потом принять участие в переносе электронов в дыхательную цепь клетки. Иными словами, эти два вещества играют важнейшую роль в клеточном дыхании.
- АТФ, то есть аденозинтрифосфат. Это вещество имеет две связи, разрыв которых даёт большое количество энергии. Этой энергией снабжаются многие жизненно важные реакции;
Также выделяются вода и углекислый газ. Давайте отразим это на нашей схеме:
Кстати, весь цикл Кребса происходит в митохондриях. Именно там, где проходит и подготовительный этап, то есть превращение пирувата в ацетил КоА. Не зря кстати митохондрии называют «энергетическая станция клетки».
Цикл Кребса как начало синтеза
Цикл Кребса удивителен тем, что он не только даёт нам ценные АТФ (энергию) и коферменты для клеточного дыхания. Если посмотрите на предыдущую схему, вы поймёте, что цикл Кребса — это продолжение процессов катаболизма. Но вместе с тем он является и первой ступенькой анаболизма. Как это возможно? Как один и тот же цикл может и разрушать, и создавать?
Оказывается, отдельные продукты реакций цикла Кребса могут частично отправляться на синтез новых сложных веществ в зависимости от потребностей организма. Например, на глюконеогенез — это синтез глюкозы из простых веществ, не являющихся углеводами.
Ещё раз:
- Реакции цикла Кребса каскадны. Они происходят одна за другой, и каждая предыдущая реакция запускает последующую;
- Продукты реакций цикла Кребса частично идут на запуск последующей реакции, а частично — на синтез новых сложных веществ.
Давайте попробуем отразить это на схеме, чтобы цикл Кребса был обозначен именно как точка пересечения распада и синтеза.
Голубыми стрелочками я отметил пути анаболизма, то есть создания новых веществ. Как видите, цикл Кребса действительно является точкой пересечения многих процессов и разрушения, и созидания.
Самое важное
- Цикл Кребса — перекрёстная точка метаболических путей. Им заканчивается катаболизм (распад), им начинается анаболизм (синтез);
- Продукты реакций Цикла Кребса частично идут для запуска следующей реакции цикла, а частично отправляются на создание новых сложных веществ;
- Цикл Кребса образует коферменты НАДH и ФАДН2, которые переносят электроны для клеточного дыхания, а также энергию в виде АТФ;
- Цикл Кребса происходит в митохондриях клеток.
Источник
Представьте себе, что все белки, жиры и углеводы, которые мы получаем с приемом пищи, распадаются на маленькие пазлы, которые собираются вновь только в одном определенном участке нашего организма – митохондриях. Там же происходит сортировка и параллельное протекание реакций созидания и разрушения. Разберемся подробнее:
Что такое цикл Кребса
Цикл Кребса – это цепочка химических реакций, происходящих в митохондриях каждой клетки нашего тела, которая называется циклом потому, что продолжается непрерывно. Она же является и общим конечным путем окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе распада большая часть органических молекул, играющих роль «клеточного топлива» или «субстратов окисления»: углеводов, жирных кислот и аминокислот.
Функции цикла Кребса:
- анаболическая (синтез новых органических веществ);
- энергетическая (питание организма)
- катаболическая (превращение некоторых веществ в катализаторы)
- транспортная (транспортировка водорода, участвующего в дыхании клеток).
Атомы водорода, высвобождающиеся в окислительно-восстановительных реакциях, доставляются в цепь переноса электронов при участии НАД- и ФАД- зависимых дегидрогеназ, в результате чего происходит образование 12 высокоэнергетических фосфатных связей: синтез 12 молекул аденозинтрифосфорной кислоты (АТФ) из аденозиндифосфорной кислоты (АДФ).
Как работает цикл Кребса?
В организме аминокислоты, жирные кислоты и пируват образуют ацетил-КоА.
Когда ацетил-КоА попадает в митохондриальный матрикс, он связывается с молекулой оксалацетата и превращается в лимонную кислоту (цитрат). Цитрат, в свою очередь, под действием фермента аконитазы превращается в цис-аконитат, оставляя молекулу воды.
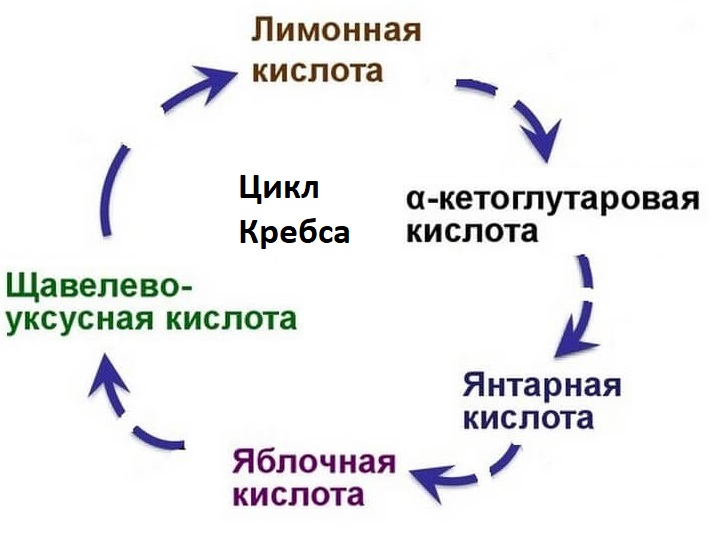
В свою очередь цис-аконитат превращается в изоцитрат под действием фермента изоцитратдегидрогеназы. Изоцитрат превращается в альфа-кетоглутарат под действием изоцитратдегидрогеназы.
Альфа-кетоглутарат превращается в сукцинил-КоА альфа-кетоглутаратдегидрогеназой и добавлением ацетил-КоА. Он подвергается сукцинату под действием сукцинат-тиокиназы. Сукцинатдегидрогеназа превращает его в фумарат. Фумарат превращается в L-малат через фумаразу. L-малат под действием фермента малатдегидрогеназы восстанавливает оксалацетат, который может снова вступать в реакцию с молекулой ацетил-КоА и повторять цикл.
Результатом этого цикла является образование CO2 и водорода, а также воды. Ионы на выходе из процесса участвуют в ресинтезе АТФ, что помогает организму восстановить еще один источник энергии – трифосфат аденозина.
Стадии цикла Кребса
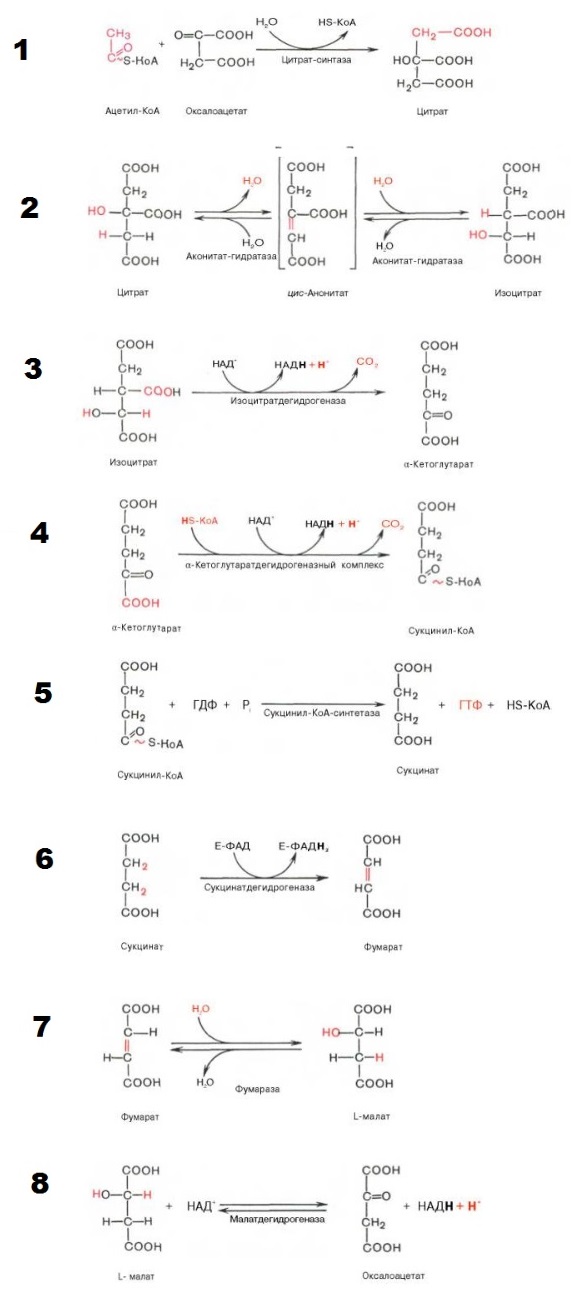
Окисление ацетильного остатка происходит в несколько стадий, образующих циклический процесс из 8 основных этапов:
Основные этапы цикла Кребса | |
I этап | Конденсация ацетил-КоА и оксалоацетата с образованием цитрата. |
Происходит реакция отщепление карбоксильной группы аминокислот, в процессе которой образуется ацетил-КоА *он выполняет функцию транспортировки углерода в различных обменных процессах. При соединении с молекулой щавелевой кислоты получается цитрат *фигурирует в буферных обменах. На данном этапе кофермент А полностью высвобождается, и получаем молекулу воды. Данная реакция необратима. | |
II этап | Превращение цитрата в изоцитрат. |
Дегидрирование (отщепление молекул воды) от цитрата, в результате которого получается цис-аконитат. И присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат) | |
III этап | Превращение изоцитрата в а-кетоглутарат. |
Изолимонная кислота(изоцитрат) дегидрируется в присутствии НАД- зависимой изо-цитратдегидрогеназы.На выходе получаем альфа-кетоглутарат. *Альфа-кетоглутарат участвует в регуляции всасывания аминокислот, нормализует метаболизм и положительно влияет на антистрессорные процессы. Также образуется NADH (аллостерический фермент) | |
IV этап | Окисление α-кетоглутарата до сукцинил-КоА |
Окислительное декарбоксилирование а-кетоглутарата с образованием сукцинил-КоА – тиоэфира, содержащего высокоэнергетическую фосфатную связь. | |
V этап | Превращение сукцинил-КоА в сукцинат. |
Пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Так же происходит образование высокоэргической фосфатной связи ГТФ за счет тиоэфирной связи сукцинил-КоА. | |
VI этап | Дегидрогенирование сукцината. Образование фумарата. |
Образовавшийся сукцинат превращается в фумарат под действием фермента сукцинат-дегидрогеназы. Единственная дегидрогеназная реакция цикла Кребса, в ходе которой осуществляется прямой перенос водорода с субстрата на флавопротеин без участия НАД+. | |
VII этап | Образование малата из фумарата. |
Под влиянием фермента фумаратгидратазы (фумаразы). Образовавшаяся при этом фумаровая кислота гидратируется, продуктом реакции является L-яблочная кислота (L-малат). | |
VIII этап | Превращение малата в оксалоацетат. |
Под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат. Происходит полное «сгорание» одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА. А коферменты (НАД+ и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. | |
Реакции цикла Кребса по стадиям
Для облегчения запоминания ферментативных реакций цикла:
ЩУКа съела ацетат, получается цитрат
через цис-аконитат будет он изоцитрат
водороды отдав НАД, он теряет СО2
этому безмерно рад альфа-кетоглутарат
окисление грядёт: НАД похитит водород
В1 и липоат с коэнзимом А спешат,
отбирают СО2, а энергия едва
в сукциниле появилась сразу ГТФ родилась
и остался сукцинат. вот добрался он до ФАДа,
водороды тому надо водороды потеряв,
стал он просто фумарат. фумарат воды напился,
и в малат он превратился тут к малату НАД пришёл,
водороды приобрёл ЩУКа снова объявилась
и тихонько затаилась Караулить ацетат…
Источник

Îäíèì èç óñëîâèé äëÿ æèçíåñïîñîáíîñòè ñëîæíîãî ìíîãîêëåòî÷íîãî îðãàíèçìà ÿâëÿåòñÿ ñòîéêàÿ è îäíîâðåìåííî ïëàñòè÷íàÿ ñèñòåìà ìåæ- è âíóòðèêëåòî÷íûõ êîììóíèêàöèé. Âçàèìîäåéñòâèÿ êëåòîê ìåæäó ñîáîé ñîñòîÿò â îáìåíå àäãåçèâíûìè, ãîðìîíàëüíûìè èëè ýëåêòðè÷åñêèìè ñèãíàëàìè. Íà áîëüøèíñòâî ïîñòóïàþùèõ â êëåòêó ñèãíàëîâ îíà ðåàãèðóåò íå íàïðÿìóþ: ìåæäó ïîñòóïàþùèì ðàçäðàæèòåëåì è ñïåöèôè÷åñêîé ðåàêöèåé êëåòêè ëåæèò öåëûé âíóòðèêëåòî÷íûé êàñêàä ñèãíàëüíûõ ìîëåêóë, ïðåäñòàâëÿþùèõ ñîáîé ïóòü áèîõèìè÷åñêèõ ïðåâðàùåíèé. Çàäà÷à òàêèõ áèîõèìè÷åñêèõ ïóòåé óñèëèòü èëè îñëàáèòü ïåðåäàâàåìûé êëåòêå ñèãíàë è ïåðåâåñòè åãî â òàêóþ ôîðìó, ÷òîáû ïîçâîëèòü ðåàëèçîâàòüñÿ îòâåòíûì ðåàêöèÿì.
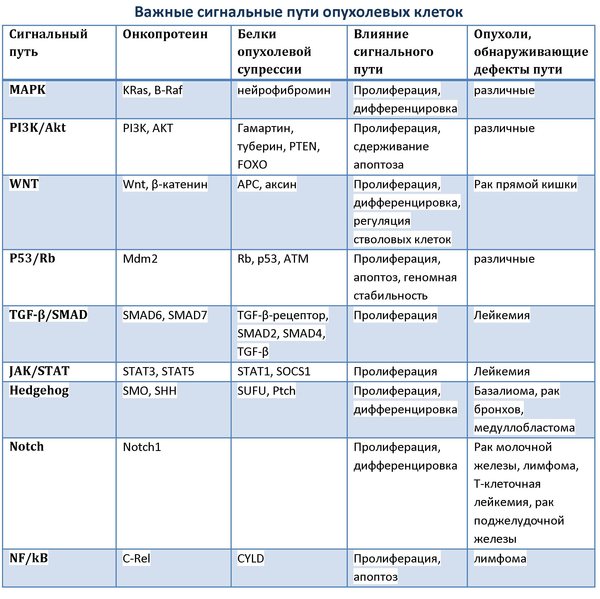
Êàê âîçíèêíîâåíèå îïóõîëåâûõ êëåòîê, òàê è ðàçëè÷èÿ ìåæäó êëåòêàìè íîðìàëüíîé è îïóõîëåâîé òêàíè áåðóò ñâî¸ íà÷àëî ñðåäè áàçîâûõ ìîëåêóëÿðíûõ ìåõàíèçìîâ. Ïîÿâëåíèå êàæäîé îïóõîëè îñíîâûâàåòñÿ íà èçìåíåíèè àêòèâíîñòè, ñâîéñòâ è âíóòðèêëåòî÷íîé êîíöåíòðàöèè îíêîïðîòåèíîâ è áåëêîâ îïóõîëåâîé ñóïðåññèè. Îäíàêî íå ñàìè áåëêè îïóõîëè êàê òàêîâûå, à èìåííî èõ áèîõèìè÷åñêîå âçàèìîäåéñòâèå è âçàèìîñâÿçè, îïîñðåäóþùèå ïåðåäà÷ó ðàçëè÷íûõ ñèãíàëîâ, ëåæàò â îñíîâå çàðîæäåíèÿ îïóõîëåâîé êëåòêè. Êîìïëåêñíàÿ âçàèìîîðãàíèçàöèÿ âñåõ çàäåéñòâîâàííûõ â ñèãíàëüíûõ ïóòÿõ áåëêîâ ôîðìèðóåò ñåòü âíóòðèêëåòî÷íîé ïåðåäà÷è ñèãíàëîâ, êîòîðàÿ ïîõîæà íà ìíîæåñòâî ñîòåí ñõîäÿùèõñÿ è âíîâü ðàçâåòâëÿþùèõñÿ, ïåðåñåêàþùèõñÿ ïóòåé, ïåðåêëþ÷àþùèõñÿ íà ðàçëè÷íûõ áåëêàõ, êîòîðûå óñëîâíî íàïîìèíàþò ñòàíöèè äëÿ ïåðåñàäîê â ìåòðî. Âñÿ ýòà ñåòü ïðåäñòàâëÿåò ñîáîé ñîâñåì íå áåñïîðÿäî÷íûé êëóáîê, êàê ìîæåò ïîêàçàòüñÿ ïîíà÷àëó, è â íåé ìîæíî âûäåëèòü íåñêîëüêî ïóòåé, êîòîðûå â êëåòêàõ áîëüøîãî êîëè÷åñòâà îïóõîëåé äåôåêòíû è îáíàðóæèâàþò îòêëîíåíèÿ îò íîðìû.
Îäíèì èç íàèáîëåå õîðîøî èçó÷åííûõ ïóòåé ÿâëÿåòñÿ ñèãíàëüíûé ïóòü MAPK (mitogen activated protein kinase), êîòîðûé íà÷èíàåòñÿ ñ àêòèâàöèè òðàíñìåìáðàííûõ ðåöåïòîðîâ êëåòêè è ïîñðåäñòâîì ó÷àñòèÿ öèòîïëàçìàòè÷åñêèõ è ÿäåðíûõ ïðîòåèíîâ ïðèíèìàåò ñóùåñòâåííîå ó÷àñòèå â ðåãóëÿöèè ãåííîé ýêñïðåññèè. Êîíå÷íî, ñòîèò îòìåòèòü, ÷òî ìíîãèå ñèãíàëüíûå ïóòè äîâîëüíî ñëîæíî ðàçëè÷àòü êàê ôóíêöèîíàëüíî, òàê è áèîõèìè÷åñêè, è ìåæäó ìíîãèìè èç íèõ çà÷àñòóþ ñóùåñòâóþò ïðÿìûå àêòèâèðóþùèå è/èëè óãíåòàþùèå ñâÿçè. Ïðèìåðîì òîìó ìîæåò ñëóæèòü ñåòü p53/Rb, îáúåäèíÿþùàÿ âàæíûå ñèãíàëüíûå ïóòè, êîòîðûå ðåãóëèðóþò ïðîöåññû êëåòî÷íîãî äåëåíèÿ, àïîïòîçà è ðåïàðàöèè ÄÍÊ.
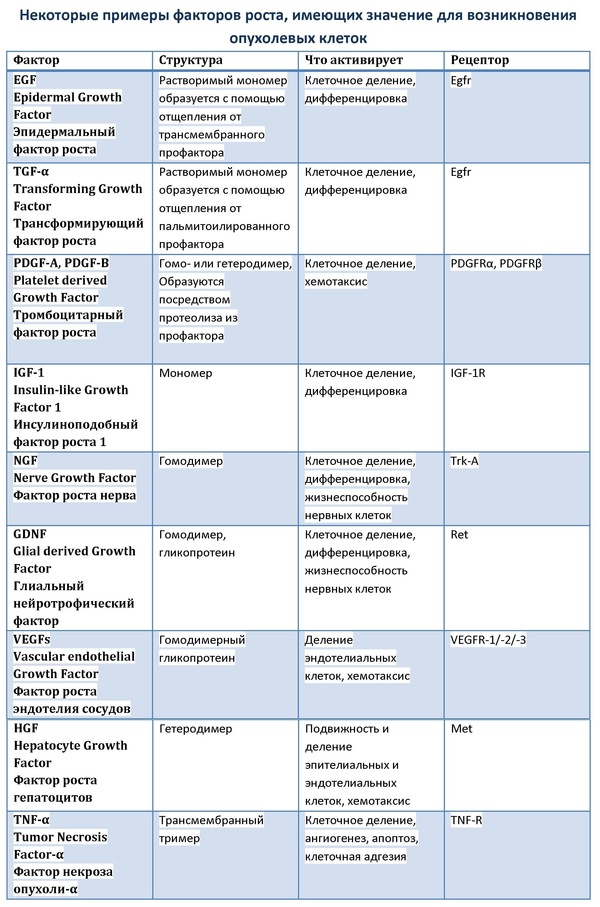
Íàøà öåëü ñåãîäíÿ óïðîù¸ííî ðàññìîòðåòü ëèøü íåêîòîðûå ïóòè, ÷òîáû ñîñòàâèòü îáùåå ïðåäñòàâëåíèå îá èõ ôóíêöèîíèðîâàíèè è ïðåäíàçíà÷åíèè. Ñèãíàëüíûé ïóòü ñîñòîèò îáû÷íî èç ôàêòîðîâ, ðåöåïòîðîâ, àäàïòîðíûõ áåëêîâ, ôåðìåíòîâ, ìîëåêóë âòîðè÷íûõ ìåññåíäæåðîâ. Êàñêàä ÌÀÐÊ íå ÿâëÿåòñÿ èñêëþ÷åíèåì. Êàê ïðàâèëî, äàííûé ñèãíàëüíûé ïóòü çàïóñêàåòñÿ ïîñðåäñòâîì ñâÿçûâàíèÿ ôàêòîðîâ ðîñòà íà ñïåöèôè÷åñêèõ ðåöåïòîðàõ ñ ïîñëåäóþùåé äèìåðèçàöèåé ðåöåïòîðîâ. Ðåöåïòîðû, ñ êîòîðûõ íà÷èíàåòñÿ ïóòü ÌÀÐÊ, îòíîñÿòñÿ ê ðåöåïòîðíûì òèðîçèíêèíàçàì (receptor tyrosine kinases, RTK). Ñòîèò îòìåòèòü, ÷òî ñî ñâÿçûâàíèÿ èìåííî ñ RTK íà÷èíàåòñÿ áîëüøîå êîëè÷åñòâî öåïî÷åê ðåàêöèé, ïðîâîäÿùèõ ê öåëåâûì ñòðóêòóðàì êëåòêè ñèãíàë ïîñëå îáðàçîâàíèÿ êîìïëåêñà ôàêòîðà ðîñòà ñ ñîîòâåòñòâóþùèì ðåöåïòîðîì.  çàâèñèìîñòè îò ïîäòèïà ðåöåïòîðà è çàäåéñòâîâàííîãî àäàïòîðíîãî áåëêà àêòèâèðóþòñÿ òå èëè èíûå ïóòè.
Ðåöåïòîðíûå òèðîçèíêèíàçû: ïðèíöèïû óñòðîéñòâà è ôóíêöèîíèðîâàíèÿ
Âñå RTK âêëþ÷àþò â ñåáÿ ãëèêîçèëèðîâàííûé âíåêëåòî÷íûé (ýêñòðàöåëëþëÿðíûé) äîìåí, íà êîòîðîì ñâÿçûâàåòñÿ ëèãàíä, ïðîíèçûâàþùèé ìåìáðàíó ãèäðîôîáíûé äîìåí è âíóòðèêëåòî÷íóþ (èíòðàöåëëþëÿðíóþ) ÷àñòü ñ þêñòàìåìáðàííûì äîìåíîì è îäíèì èëè äâóìÿ êàòàëèòè÷åñêèìè òèðîçèíêèíàçíûìè äîìåíàìè. Êàòàëèòè÷åñêèå äîìåíû ñîñòîÿò èç àìèíîòåðìèíàëüíîãî ÀÒÔ-ñâÿçûâàþùåãî ðåãèîíà è êàðáîêñèòåðìèíàëüíîãî ôîñôîòðàíñôåðàçíîãî äîìåíà. Ìíîãèå ãåíû ðåöåïòîðíûõ òèðîçèíêèíàç ÿâëÿþòñÿ ãîìîëîãàìè âèðóñíûõ îíêîãåíîâ. Ïî ñòðóêòóðíûì õàðàêòåðèñòèêàì âíåêëåòî÷íûõ äîìåíîâ RTK ìîæíî ðàçäåëèòü íà íåñêîëüêî êëàññîâ. Ê ïðèìåðó, ïðåäñòàâèòåëè ñåìåéñòâ PDGFR (ðåöåïòîð òðîìáîöèòàðíîãî ôàêòîðà ðîñòà), FGFR (ðåöåïòîð ôàêòîðà ðîñòà ôèáðîáëàñòîâ), VEGFR-1/-2 (ðåöåïòîðà ôàêòîðà ðîñòà ýíäîòåëèÿ ñîñóäîâ) îáëàäàþò ñîîòâåòñòâåííî ïÿòüþ, òðåìÿ è ñåìüþ Ig-ïîäîáíûìè äîìåíàìè. Îäíàêî, íåñìîòðÿ íà ìíîãîîáðàçèå êëàññîâ ðåöåïòîðíûõ òèðîçèíêèíàç, ìåõàíèçì èõ àêòèâàöèè ïðàêòè÷åñêè îäèíàêîâ. Ñîãëàñíî ýòîé ìîäåëè, â íåñâÿçàííîì ñîñòîÿíèè ìåæäó àêòèâíûìè äèìåðàìè è íåàêòèâíûìè ìîíîìåðàìè ðåöåïòîðà ïîñòîÿííî ïîääåðæèâàåòñÿ ðàâíîâåñèå. Ïðèñîåäèíåíèå ëèãàíäà âåä¸ò ê äèìåðèçàöèè è ñìåùàåò ðàâíîâåñèå â ñòîðîíó îáðàçîâàíèÿ àêòèâíîé ôîðìû ðåöåïòîðà. Îáðàçîâàíèå àêòèâíûõ äèìåðîâ ìîæåò áûòü èíèöèèðîâàíî íàïðÿìóþ ëèãàíäàìè, ñâÿçûâàþùèìèñÿ ñðàçó ñ äâóìÿ ìîíîìåðàìè, êàê, íàïðèìåð, EGF (epidermal growth factor) ñïîñîáñòâóåò äèìåðèçàöèè ñâîåãî ðåöåïòîðà Egfr. Òàêæå ñâÿçûâàíèå ñ ëèãàíäîì ìîæåò âûçûâàòü èçìåíåíèå êîíôîðìàöèè âíåêëåòî÷íîãî äîìåíà, ÷òî âåäåò ê ýêñïîçèöèè ñàéòîâ ñâÿçûâàíèÿ, êàê íàïðèìåð, SCF (stem cell factor) âûçûâàåò äèìåðèçàöèþ Kit-ðåöåïòîðîâ. Èòîãîì ïðîöåññà äèìåðèçàöèè ÿâëÿåòñÿ ñáëèæåíèå âíóòðèêëåòî÷íûõ äîìåíîâ äðóã ñ äðóãîì, âñëåäñòâèå ÷åãî íàñòóïàåò ïðåõîäÿùàÿ àêòèâàöèÿ âíóòðåííåé òèðîçèíêèíàçíîé àêòèâíîñòè êàòàëèòè÷åñêèõ äîìåíîâ, ÷òî ïðèâîäèò ê òðàíñôîñôîðèëèðîâàíèþ ñïåöèôè÷åñêèõ îñòàòêîâ òèðîçèíà öèòîïëàçìàòè÷åñêîãî äîìåíà. Ñ ôîñôîðèëèðîâàííûì ðåöåïòîðîì ìîãóò ñâÿçûâàòüñÿ áåëêè ñ SH2-äîìåíàìè. Êîìïëåêñ ñâÿçàííîãî ñ òàêèì áåëêîì ðåöåïòîðà ìîæåò îïîñðåäîâàòü àêòèâàöèþ, íàïðèìåð, êàêîãî-ëèáî ôåðìåíòà èëè èçìåíåíèå ðåàêöèîííîé ñïîñîáíîñòè áåëêà.  îòñóòñòâèå ëèãàíäîâ RTK ïðåäñòàâëÿþò ñîáîé ìîíîìåðíûå ïîëèïåïòèäíûå öåïî÷êè (èñêëþ÷åíèå ñåìåéñòâî ðåöåïòîðîâ èíñóëèíà, êîòîðûå ñîñòîÿò èç 4 ïåïòèäíûõ öåïåé, ñîåäèí¸ííûõ äèñóëüôèäíûìè ìîñòèêàìè).
Êëàññè÷åñêàÿ ïîñëåäîâàòåëüíîñòü ðàçâèòèÿ ïóòè ÌÀÐÊ
Ïîñëå íåáîëüøîãî îòâëå÷åíèÿ íà îïèñàíèå ðåöåïòîðíîãî âçàèìîäåéñòâèÿ, âåðíåìñÿ ê ïóòè ÌÀÐÊ. Àêòèâàöèÿ RTK, ïðåäâàðÿþùàÿ ðàçâèòèå ñîáûòèé âíóòðè êëåòêè, ïðèâîäèò ê ñâÿçûâàíèþ SH2-äîìåíà àäàïòîðíîãî áåëêà GRB2 (Growth Factor Receptor bound 2) íà ôîñôîðèëèðîâàííîì îñòàòêå òèðîçèíà àêòèâèðîâàííîé RTK. Ïîìèìî SH2-äîìåíà, GRB2 ñîäåðæèò òàêæå äâà SH3-äîìåíà, èìåþùèõ ñðîäñòâî ê âçàèìîäåéñòâèþ ñ áîãàòûìè ïðîëèíîì ó÷àñòêàìè áåëêà, êîòîðûé ãîìîëîãè÷åí áåëêó ïëîäîâîé ìóøêè Drosophila SOS (son of sevenless) è ïîòîìó ó ìëåêîïèòàþùèõ äàííûé áåëîê òàêæå èìååò íàçâàíèå SOS. Ýòîò áåëîê ÿâëÿåòñÿ ôàêòîðîì îáìåíà ãóàíèíîâûõ íóêëåîòèäîâ è îïîñðåäîâàííî (ñ ïîìîùüþ îäíîãî èç Ras-áåëêîâ: HRas, KRas, NRas) êàòàëèçèðóåò îáìåí ÃÄÔ íà ÃÒÔ. Íåîáõîäèìîñòü GRB2 äëÿ àêòèâàöèè Ras è ìèòîãåííîãî ýôôåêòà ÌÀÐÊ äîêàçàíà ýêñïåðèìåíòàëüíî. Òàêæå ñîâìåñòíàÿ èíúåêöèÿ GRB2 è HRas â ïîêîÿùèåñÿ ôèáðîáëàñòû âåäåò ê ñèíòåçó ÄÍÊ, ÷åãî íå íàáëþäàåòñÿ ïðè èíúåöèðîâàíèè îòäåëüíî GRB2 èëè HRas. Èíúåêöèÿ àíòèòåë ïðîòèâ GRB2 ïðåïÿòñòâóåò âñòóïëåíèþ êëåòîê â S-ïåðèîä êëåòî÷íîãî öèêëà è ïåðåñòðîéêå êëåòî÷íîãî öèòîñêåëåòà ïîñëå îáðàáîòêè èõ EGF èëè PDGF. GRB2 ñâÿçûâàåòñÿ ñ ïîìîùüþ ñâîåãî SH2-äîìåíà íå òîëüêî ñ ôîñôîòèðîçèíîì àêòèâèðîâàííîé ðåöåïòîðíîé òèðîçèíêèíàçû, íî è ñ ôîñôîòèðîçèíîì àäàïòîðíîãî áåëêà SHC, ôîñôîòèðîçèíôîñôàòàçîé SHP-2 è õèìåðíûì áåëêîì BCR-Abl.
 ÃÒÔ-ñâÿçàííîé ôîðìå áåëêè Ras ñïîñîáíû àêòèâèðîâàòü è äðóãèå ïðîòåèíû. Ñðåäè âàæíåéøèõ ýôôåêòîðîâ Ras ìîæíî âûäåëèòü ôîñôàòèäèëèíîçèòîë-3-êèíàçó (PI3K), ôàêòîð îáìåíà Ral (RalGEF) è ôîñôîëèïàçó Ñ (PLC). Òàêèì îáðàçîì, ñ ïîìîùüþ Ras-ïðîòåèíîâ ïðîèñõîäèò àêòèâàöèÿ íåñêîëüêèõ ñèãíàëüíûõ ïóòåé, íåðàçðûâíî ñâÿçàííûõ ìåæäó ñîáîé. Ras ðåãóëèðóþò íå òîëüêî ïðîëèôåðàöèþ êëåòîê, íî è êëåòî÷íóþ ìèãðàöèþ, äèôôåðåíöèðîâêó, àíãèîãåíåç è êîíòðîëèðóþò ïðåïÿòñòâóþùèå àïîïòîçó ìåõàíèçìû.
Ñàìûì èçâåñòíûì ýôôåêòîðîì Ras ÿâëÿåòñÿ ñåðèí-/òðåîíèíêèíàçà B-Raf, êîòîðàÿ âåäåò ê çàïóñêó êëàññè÷åñêîãî ïóòè ÌÀÐÊ. Ðåøàþùèì ìîìåíòîì äëÿ àêòèâàöèè Â-Raf-êèíàçû ÿâëÿåòñÿ íå ñâÿçûâàíèå íà Ras-GTP, à ïåðåìåùåíèå íà âíóòðåííþþ ïîâåðõíîñòü ïëàçìàòè÷åñêîé ìåìáðàíû.  ýêñïåðèìåíòàëüíîì èññëåäîâàíèè åù¸ 1994 ãîäà áûëî ïîäòâåðæäåíî, ÷òî åñëè çàÿêîðèòü Â-Raf-ïðîòåèí â ìåìáðàíå ñ ïîìîùüþ ëèïèäíîãî «ÿêîðÿ», òî îí áóäåò àêòèâíûì è áåç âçàèìîñâÿçè ñ Ras-GTP. Ïîýòîìó ñ÷èòàåòñÿ, ÷òî Â-Raf âçàèìîäåéñòâóåò íà âíóòðåííåé ïîâåðõíîñòè ìåìáðàíû ñ êàêèì-ëèáî ôàêòîðîì áåëêîâîé èëè ëèïèäíîé ïðèðîäû (êàêèì ïîêà íåèçâåñòíî), ÷òî è ñòàíîâèòñÿ óñëîâèåì äëÿ àêòèâàöèè áåëêà. Äëÿ çàâåðøåíèÿ àêòèâèðîâàíèÿ áåëîê äîëæåí áûòü òàêæå ôîñôîðèëèðîâàí, çà ÷òî, âåðîÿòíî, îòâå÷àåò Src-êèíàçà. B-Raf-êèíàçà ôîñôîðèëèðóåò è ïðèâîäèò â àêòèâíîå ñîñòîÿíèå êèíàçû ÌÅÊ 1 è 2, êîòîðûå, â ñâîþ î÷åðåäü, íåîáõîäèìû äëÿ àêòèâèðîâàíèÿ êèíàç ERK 1 è 2. ERK 1 è 2, áóäó÷è àêòèâíûìè, ñïîñîáñòâóþò ôîñôîðèëèðîâàíèþ è àêòèâàöèè ìíîãî÷èñëåííûõ ôàêòîðîâ òðàíñêðèïöèè, êàê, íàïðèìåð, Elk-1, fos, Myk. Ôàêòîðû òðàíñêðèïöèè ñâÿçûâàþòñÿ ñî ñïåöèôè÷åñêèìè ïîâòîðàìè íà ìîëåêóëå ÄÍÊ è èíèöèèðóþò ïðîöåññ òðàíñêðèïöèè íåîáõîäèìûõ ãåíîâ, íàïðèìåð, CCND1, FOS. Äàëåå ñèíòåçèðîâàííûå â õîäå òðàíñëÿöèè ïðîäóêòû äàííûõ ãåíîâ îïîñðåäóþò ðàçâèòèå áèîëîãè÷åñêîé ðåàêöèè êëåòêè íà àêòèâèðîâàíèå ñèãíàëüíîãî ïóòè.
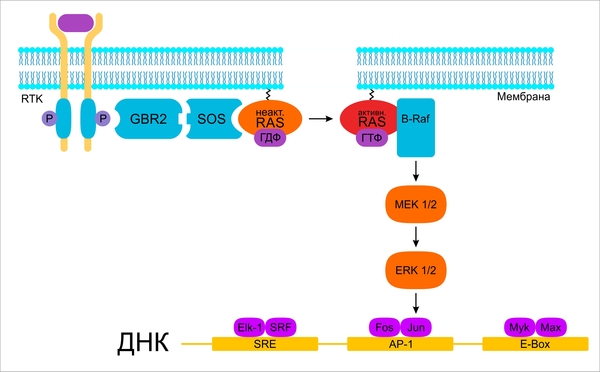
Ñõåìà êëàññè÷åñêîãî ñèãíàëüíîãî ïóòè ÌÀÐÊ.
ÌÀÐÊ â êëåòêàõ îïóõîëè
Óñèëåííàÿ àêòèâàöèÿ ïóòè ÌÀÐÊ ìîæåò áûòü âûçâàíà ìóòàöèÿìè â ãåíàõ RTK, à òàêæå HRas, KRas, NRas, BRAF1, NF1. Îäíàêî àêòèâèðóþùèå ìóòàöèè â ãåíàõ, ÷üè ïðîäóêòû ïðèíèìàþò ó÷àñòèå â ïðîâåäåíèè ñèãíàëà âíóòðü êëåòêè, ìîãóò áûòü ñêîìïåíñèðîâàíû, ïîñêîëüêó, êàê ïðàâèëî, åñëè îäèí èç ó÷àñòêîâ ñèãíàëüíîãî ïóòè èçìåí¸í âñëåäñòâèå ìóòàöèè, òî îñòàëüíûå îñòàþòñÿ íåòðîíóòûìè.  êà÷åñòâå ïðèìåðà ìîæíî ïðèâåñòè èññëåäîâàíèå êëåòîê ïàïèëëÿðíîãî ðàêà ÙÆ. Äâå òðåòè êàðöèíîì íåñëè àêòèâèðóþùèå ìóòàöèè èëè â ãåíå ðåöåïòîðíîé òèðîçèíêèíàçû RET, èëè KRAS, èëè BRAF1. È òîëüêî â î÷åíü íåáîëüøîì ïðîöåíòå îïóõîëåé áûëè îáíàðóæåíû ìóòàöèè â äâóõ ðàçëè÷íûõ ãåíàõ ñèãíàëüíîãî ïóòè. Ïî÷åìó â îäíîé îïóõîëè ïîâðåæäåí îäèí ãåí, à â äðóãîé èíîé ãåí îäíîãî è òîãî æå ñèãíàëüíîãî ïóòè, ïîêà íå ñîâñåì ïîíÿòíî. Îäíî èç ïðåäïîëîæåíèé îïèðàåòñÿ íà èåðàðõèþ â ïðåäåëàõ ïóòè è ìóòàöèé. ÌÀÐÊ ìîæåò ðàçâåòâëÿòüñÿ íà íåñêîëüêèõ óðîâíÿõ, è ÷åì âûøå ïî èåðàðõèè ñèãíàëüíîãî ïóòè ëåæèò èçìåí¸ííûé áåëîê, òåì áîëåå âåðîÿòíî ïîñëåäóþùåå îòêëîíåíèå. Ñëåäóÿ ýòîìó ïðåäïîëîæåíèþ, ìóòàöèÿ â ãåíàõ Ras áóäåò èìåòü áîëåå îáøèðíûå è âûðàæåííûå ïîñëåäñòâèÿ äëÿ êëåòêè, ÷åì, íàïðèìåð, ìóòàöèÿ â ãåíå BRAF. Ñëåäîâàòåëüíî, ìóòàöèè â ãåíàõ Ras áîëåå «âûãîäíû» ïîòåíöèàëüíîé îïóõîëåâîé êëåòêå è âñòðå÷àþòñÿ ÷àùå, ÷åì ìóòàöèè íà ôóíêöèîíàëüíî íèæåðàñïîëîæåííûõ óðîâíÿõ.
Áåëêè Ras çàêðåïëåíû íà êëåòî÷íîé ìåìáðàíå ñ âíóòðåííåé ñòîðîíû ïîñðåäñòâîì æèðíîé êèñëîòû, êîâàëåíòíî ñâÿçàííîé ñ êàðáîêñèòåðìèíàëüíûì êîíöîì áåëêà. È ýòî ñåìåéñòâî áåëêîâ ìîæíî ñðàâíèòü ñ ìîëåêóëÿðíûì ïåðåêëþ÷àòåëåì ÌÀÐÊ.  ÃÄÔ-ñâÿçàííîì ñîñòîÿíèè áåëîê íåàêòèâåí, è äàëüíåéøàÿ ïåðåäà÷à ñèãíàëà íå îñóùåñòâëÿåòñÿ. Îáìåí ÃÄÔ íà ÃÒÔ è ñîïðÿæ¸ííûé ñ ýòèì ïåðåõîä íåàêòèâíîé ôîðìû Ras â àêòèâíóþ êàòàëèçèðóåòñÿ SOS (SOS1, SOS2) ôåðìåíòîì, îòíîñÿùèìñÿ ê ãðóïïå ôàêòîðîâ îáìåíà (GEF Guanine Exchange Factors). Àêòèâíàÿ ôîðìà Ras ìîæåò ïðîÿâëÿòü ñîáñòâåííóþ íåâûñîêóþ ÃÒÔ-àçíóþ àêòèâíîñòü è ãèäðîëèçîâàòü ñâÿçàííûé ÃÒÔ äî ÃÄÔ, òåì ñàìûì ñàìîèíàêòèâèðóÿñü. Ýòîò ïðîöåññ äîâîëüíî äëèòåëåí, íî îí ìíîãîêðàòíî óñêîðÿåòñÿ ÃÒÔ-àçà-àêòèâèðóþùèìè áåëêàìè, íàïðèìåð, p120GAP (p120 GTPase activating Protein) è íåéðîôèáðîìèíîì (NF1GAP). Ìóòàöèè â ãåíàõ îíêîïðîòåèíîâ Ras âåäóò ê ïðåäîòâðàùåíèþ ðåàêöèè ãèäðîëèçà, âñëåäñòâèå ÷åãî ñèãíàëüíûé ïóòü äëèòåëüíîå âðåìÿ îñòà¸òñÿ àêòèâíûì.
Ñåìåéñòâî Rho-ÃÒÔàç è èõ ó÷àñòèå â ðàçâèòèè îïóõîëåâîãî ïðîöåññà
Íàðÿäó ñ Ras ñóùåñòâóþò è äðóãèå ÃÒÔ-ñâÿçûâàþùèå áåëêè, êîòîðûå ñ ïîìîùüþ ïîäîáíûõ áèîõèìè÷åñêèõ ìåõàíèçìîâ àêòèâèðóþòñÿ èëè èíàêòèâèðóþòñÿ è èìåþò ðàçëè÷íûå çàäà÷è. È ñðåäè ìàëåíüêèõ ÃÒÔ-àç õîòåëîñü áû óïîìÿíóòü ñåìåéñòâî Rho, ïðåäñòàâèòåëè êîòîðîãî èãðàþò êëþ÷åâóþ ðåãóëÿòîðíóþ ðîëü â ñèãíàëüíîé ïåðåäà÷å îò öèòîêèíîâûõ ðåöåïòîðîâ, à òàêæå â îðãàíèçàöèè àêòèíîâîãî öèòîñêåëåòà êëåòêè è ìèêðîòðóáî÷åê.  ðåàëèçàöèè ïðîöåññà ìåòàñòàçèðîâàíèÿ öèòîñêåëåò èìååò áîëüøîå çíà÷åíèå, ïîñêîëüêó ïîìèìî ìåõàíè÷åñêîé ñòàáèëèçàöèè êëåòî÷íîé àðõèòåêòóðû, ñåíñîðíûå ôóíêöèè è òðàíñïîðò âíóòðè êëåòêè íåîáõîäèìû äëÿ èíâàçèâíîé àêòèâíîñòè êëåòêè. Ñ ïîìîùüþ äèíàìè÷åñêîé ñáîðêè è ðàçáîðêè ñòðóêòóð öèòîñêåëåòà êëåòêà ìîæåò àêòèâíî ïåðåäâèãàòüñÿ. Rho ÃÒÔ-àçû àêòèâíû â ÃÒÔ-ñâÿçàííîì ñîñòîÿíèè, à â ÃÄÔ-ñâÿçàííîé ôîðìå, ñîîòâåòñòâåííî, íå ïðîÿâëÿþò ôóíêöèîíàëüíîé àêòèâíîñòè. GEF-áåëêè êàòàëèçèðóþò îáìåí ÃÄÔ íà ÃÒÔ, îïîñðåäóÿ àêòèâíîñòü Rho ÃÒÔ-àç, à áåëêè GAP ñòèìóëèðóþò èíàêòèâàöèþ Rho-áåëêîâ. Àêòèâíûå Rho ÃÒÔ-àçû ðàñïîëàãàþòñÿ íà êëåòî÷íîé ìåìáðàíå, ãäå, ñâÿçûâàÿ ñïåöèôè÷åñêèå ýôôåêòîðíûå áåëêè, îíè îáåñïå÷èâàþò ïîñëåäóþùóþ òðàíñäóêöèþ ñèãíàëà, âûçûâàÿ ïåðåãðóïïèðîâêó öèòîñêåëåòà. Òîò ôàêò, ÷òî íà ñåãîäíÿøíèé äåíü èäåíòèôèöèðîâàíî îêîëî 80-òè òèïîâ GEF è 70-òè GAP, ÷òî êîëè÷åñòâåííî ïðåâîñõîäèò Rho ÃÒÔ-àçû, êîèõ èìååòñÿ îêîëî 20-òè, ìîæåò ãîâîðèòü î íåîáõîäèìîñòè ñòðîãîãî êîíòðîëÿ ðåãóëÿöèè ëîêàëüíîé àêòèâíîñòè Rho, ÷òîáû ïðåäîòâðàòèòü îøèáî÷íóþ ïåðåäà÷ó èñõîäíîãî ñèãíàëà.
 îïóõîëåâûõ êëåòêàõ àêòèâíîñòü Rho ÃÒÔ-àç íåíîðìàëüíî âûñîêà, ÷òî ìîæåò áûòü îáóñëîâëåíî èçìåí¸ííîé ãåííîé ýêñïðåññèåé èëè íàðóøåííîé ôóíêöèåé ðåãóëÿòîðîâ, â ìåíüøåé ñòåïåíè àêòèâèðóþùèìè ìóòàöèÿìè â ñàìèõ Rho ÃÒÔ-àçàõ. Ñðåäè RhoGAP-áåëêîâ îñîáåííî âûäåëÿþò ñåìåéñòâî DLC-áåëêîâ (deleted in liver cancer), ïîñêîëüêó èìåííî èõ èíàêòèâàöèÿ ÿâëÿåòñÿ íàèáîëåå ÷àñòûì èçìåíåíèåì Rho-ðåãóëÿòîðîâ ïðè ðàçâèòèè îïóõîëåâûõ ïðîöåññîâ. Ïðè íåêîòîðûõ òèïàõ ðàêà óòðàòà DLC1 âñòðå÷àåòñÿ íàñòîëüêî æå ÷àñòî, êàê è âûïàäåíèå îïóõîëåâîãî ñóïðåññîðà ð53. Âîîáùå â ãåíîìå ÷åëîâåêà çàêîäèðîâàíû òðè èçîôîðìû DLC, èìåþùèå öèôðîâîå îáîçíà÷åíèå îò 1 äî 3. DLC1-3 îáëàäàþò ñõîæåé ñòðóêòóðíîé îðãàíèçàöèåé è ðåãóëèðóþò àêòèâíîñòü ìàëûõ ÃÒÔ-àç RhoÀ è Cdc42. DLC1 ÿâëÿåòñÿ íàèáîëåå èçó÷åííûì ïðåäñòàâèòåëåì è áûë îòêðûò â 1998 ãîäó êàê âåðîÿòíûé îïóõîëåâûé ñóïðåññîð â êëåòêàõ ãåïàòîöåëëþëÿðíîé êàðöèíîìû ïå÷åíè ÷åëîâåêà. Ïîçæå áûëî óñòàíîâëåíî, ÷òî áåëîê DLC1 îòñóòñòâóåò â êëåòêàõ ðàçëè÷íûõ îïóõîëåâûõ îáðàçîâàíèé (ìîëî÷íîé æåëåçû, êèøå÷íèêà, ë¸ãêèõ, ïðîñòàòû) ïî ïðè÷èíå èíàêòèâàöèè ñîîòâåòñòâóþùåãî åìó ãåíà. Ïåðåñòðîéêà ýêñïðåññèè DLC1 â êëåòêàõ ðàçëè÷íûõ îïóõîëåâûõ ëèíèé, íå îáíàðóæèâàâøèõ îáðàçîâàíèå ýíäîãåííîãî DLC1, îñòàíàâëèâàëà èíòåíñèâíûé ðîñò êëåòîê è äàëüíåéøåå ðàçâèòèå îïóõîëè íà ìîäåëè ìûøè. Êðîìå òîãî, îáåñïå÷åíèå ñòàáèëüíîé ýêñïðåññèè DLC1 â êëåòêàõ ðàêà ìîëî÷íîé æåëåçû ÷åëîâåêà ïðèâåëî ê óìåíüøåíèþ ìèãðàöèè êëåòîê in vitro è ê ñîêðàùåíèþ êëåòî÷íîé èíâàçèè in vivo.
Ñîòðóäíèêàìè ëàáîðàòîðèè Èíñòèòóòà êëåòî÷íîé áèîëîãèè è èììóíîëîãèè Óíèâåðñèòåòà ãîðîäà Øòóòòãàðòà áûëè èññëåäîâàíû ïîñëåäñòâèÿ âûïàäåíèÿ áåëêà DLC1 èç ñèãíàëüíîãî ïóòè ñ ïîìîùüþ ÐÍÊ-èíòåðôåðåíöèè. Ñ ïðèìåíåíèåì êîíôîêàëüíîé èììóíîôëóîðåñöåíòíîé ìèêðîñêîïèè áûëî ïîêàçàíî íà ïðèìåðå êëåòîê ðàêà ìîëî÷íîé æåëåçû, ÷òî «âûêëþ÷åíèå» DLC1 âûçûâàåò óñèëåííîå îáðàçîâàíèå ò.í. àêòèâíîâûõ ñòðåññîâûõ âîëîêîí è ôîêàëüíûõ àäãåçèâíûõ êîíòàêòîâ. Ýòî áûëî ñîïðÿæåíî òàêæå ñ ïîâûøåííîé ìèãðàöèîííîé àêòèâíîñòüþ ðàêîâûõ êëåòîê, ÷òî áûëî çàôèêñèðîâàíî â àíàëèçå ìèãðàöèè êëåòîê ÷åðåç ìåìáðàíó, ïðîâåä¸ííîì â ñïåöèàëüíûõ Transwell-êóëüòóðàëüíûõ ïëàíøåòàõ, â êîòîðûõ ìîæíî ìîäåëèðîâàòü õåìîòàêñè÷åñêóþ ìèãðàöèþ êëåòîê ÷åðåç íåáîëüøèå ïîðû. Ìåõàíèñòè÷åñêè óâåëè÷åíèå ìèãðàöèîííîé àêòèâíîñòè ìîæåò áûòü îáúÿñíåíî óñèëåíèåì Rho-îïîñðåäîâàííîé ïåðåäà÷è ñèãíàëà êàê ñëåäñòâèå óòðàòû DLC1. À íà ïðèìåðå èñïîëüçîâàíèÿ êëåòîê ðàêà ïå÷åíè ìûøè áûëà îäíîçíà÷íî ïîäòâåðæäåíà ôóíêöèÿ DLC1 êàê îïóõîëåâîãî ñóïðåññîðà.
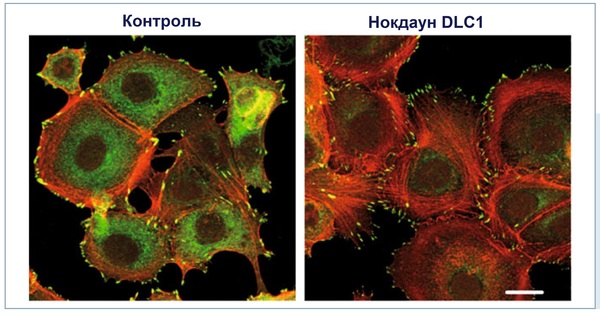
Îáðàçîâàíèå ñòðåññîâûõ âîëîêîí è ôîêàëüíûõ àäãåçèâíûõ êîíòàêòîâ ïóòåì ñíèæåíèÿ ýêñïðåññèè DLC1. Êëåòî÷íûé îòâåò íà ñíèæåííóþ ýêñïðåññèþ DLC1 â ëèíèè îïóõîëåâûõ êëåòîê MCF7 (êëåòêè ðàêà ìîëî÷íîé æåëåçû) áûë ïîëó÷åí ïîñëå ïðîâåäåíèÿ ÐÍÊ-èíòåðôåðåíöèè. Ïðåäñòàâëåííûå ôîòîãðàôèè îòðàæàþò ðåçóëüòàòû èììóíîôëóîðåñöåíòíîé ìèêðîñêîïèè. Îêðàñêà àäãåçèâíûõ êîíòàêòîâ ïðîèçâåäåíà ñ ïîìîùüþ ñïåöèôè÷åñêèõ àíòèòåë ê ïàêñèëèíó (çåëåíûé öâåò), îêðàñêà ñòðåññîâûõ âîëîêîí ñ ïîìîùüþ ôàëëîèäèíà (êðàñíûé öâåò).
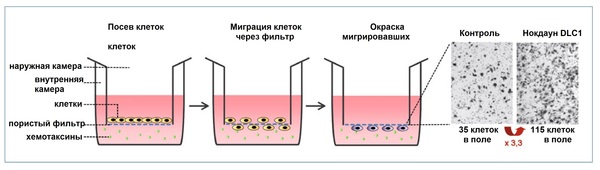
Transwell-àíàëèç äëÿ èññëåäîâàíèÿ õåìîòàêñè÷åñêîé ìèãðàöèè êëåòîê. Íîêàóòèðîâàííûå ïî DLC1 êëåòêè, à òàêæå êëåòêè êîíòðîëüíîé ãðóïïû áûëè ïîñåÿíû íà ñïåöèàëüíûé ïëàíøåò; èõ ìèãðàöèÿ ÷åðåç ïîðû ôèëüòðà èíäóöèðîâàíà íàëè÷èåì ãðàäèåíòà ñûâîðîòêè è êîëëàãåíà. Ïîñëå ýòîãî ìèãðèðîâàâøèå êëåòêè áûëè îêðàøåíû è ïîäñ÷èòàíû.
×òî æå ñ äðóãèìè ÷ëåíàìè ñåìüè DLC? Íåñìîòðÿ íà òî, èõ õàðàêòåðèñòèêà ñîñòàâëåíà åùå íå â ïîëíîé ìåðå, èõ îòñóòñòâèå îòìå÷åíî â êëåòêàõ ìíîãèõ îïóõîëåé. Òàêæå èíòåðåñåí òîò ôàêò, ÷òî, íåñìîòðÿ íà ñõîæåñòü ñòðóêòóð îòäåëüíûõ èçîôîðì DLC, èõ ôóíêöèè, òåì íå ìåíåå, íå ÿâëÿþòñÿ èçáûòî÷íûìè, à ñàìè áåëêè íå îáëàäàþò ïîëíîñòüþ âçàèìîêîìïåíñèðóþùèì ñâîéñòâîì. Îá ýòîì ñâèäåòåëüñòâóåò òî, ÷òî óòðàòà ãåíà DLC1 ïðèâîäèò ê ëåòàëüíîìó ôåíîòèïó ó ìûøåé, òîãäà êàê íîêàóòèðîâàííûå ïî ãåíó DLC2 ìûøè îñòàþòñÿ æèçíåñïîñîáíûìè.  ñâÿçè ñ ýòèì ôàêòîì äîâîëüíî çíà÷èòåëüíûìè ïðåäñòàâëÿþòñÿ ðàáîòû èññëåäîâàòåëåé èç Øòóòòãàðòà, ïîñêîëüêó â ñâîèõ ýêñïåðèìåíòàõ èì óäàëîñü òàêæå îáíàðóæèòü ñïåöèôè÷íóþ äëÿ èçîôîðìû DLC3 ôóíêöèþ, çàêëþ÷àþùóþñÿ â êîîðäèíàöèè ïðîöåññîâ âíóòðèêëåòî÷íîãî òðàíñïîðòà ìåæäó îòäåëüíûìè êîìïàðòìåíòàìè êëåòêè. Ïîìèìî ýòîãî, DLC3 îñóùåñòâëÿåò ïåðåäà÷ó ñèãíàëîâ îò ðåöåïòîðà ôàêòîðà ðîñòà EGFR (epidermal growth factor receptor) è âûïîëíÿåò ôóíêöèþ ñòàáèëèçàöèè â ïîñòîÿííûõ êëåòî÷íûõ êîíòàêòàõ. Óòðàòà ôóíêöèè DLC3 â îïóõîëåâûõ êëåòêàõ ìîæåò, ñ îäíîé ñòîðîíû, ïðèâåñòè ê óñèëåíèþ ñèãíàëîâ â êëåòêó îò ðåöåïòîðîâ ðîñòîâûõ ôàêòîðîâ è, ñ äðóãîé ñòîðîíû, ñòàòü ïðè÷èíîé îñëàáëåíèÿ êîíòàêòîâ ìåæäó ñòðóêòóðàìè ýïèòåëèàëüíûõ òêàíåé, ÷òî ñïîñîáñòâóåò êàê âîçíèêíîâåíèþ îïóõîëè, òàê è å¸ ìåòàñòàçèðîâàíèþ.
Íà ýòîì ïðèä¸òñÿ ïîäâåñòè èòîãè. Ñèãíàëüíûõ ïóòåé â êëåòêå âåëèêîå ìíîæåñòâî, ìû âçÿëè òîëüêî íåñêîëüêî è ïðîáåæàëèñü ïî îñíîâíûì ìîìåíòàì. Ó âàñ óæå íàâåðíÿêà âîçíèêëî ìíîæåñòâî âîïðîñîâ êàñàòåëüíî ïóòåé, èõ êîìïîíåíòîâ è èõ ôóíêöèîíèðîâàíèÿ â îïóõîëåâûõ êëåòêàõ.  õîäå íàïèñàíèÿ ïîñëåäóþùèõ î÷åðêîâ ïî ñèãíàëüíûì ïóòÿì áóäóò ïðèëîæåíû âñå óñèëèÿ, ÷òîáû îòâåòèòü íà âàøè âîïðîñû è ÷óòü øèðå ïðèîòêðûòü äëÿ âñåõ èíòåðåñóþùèõñÿ âíóòðåííèé ìèð êëåòêè.
Âñïîìîãàòåëüíûå ïîíÿòèÿ è àááðåâèàòóðû
Àâòîð: Àëåêñàíäðà Ñòåöåíêî
Èñòî÷íèêè:
C. Wagener, O. Müller Molekulare Onkologie: Entstehung, Progressing, klinische Aspekte, 2010
A. Nordheim, R. Knippers Molekulare Genetik, 2015
Lowenstein, Daly, Batzer et al. The SH2 and SH3 Domain-Containing Protein GRB2 Links Receptor Tyrosine Kinases to ras Signaling
Van der Geer, Hunter, Lindberg Receptor protein-tyrosine kinases and their signal transduction pathways
Wittinghofer, Pai The structure of Ras protein: a model for a universal molecular switch
Noll et al. Rho-Signalgebung in der Tumorentstehung und -progression, BioSpektrum, 04.2016
Braun, Hendrick, Eisker, Schmid et al. The Rho-specific GAP protein DLC3 coordinates endocytic membrane trafficking
Braun, Olayioye Rho regulation: DLC proteins in space and time
Источник